DESARROLLO Y DIFERENCIACIÓN EN EL EMBRIÓN
Regulación Genética
Germán Arellano Cruz
________________________________________________________
La embriología es el estudio del desarrollo del huevo, desde la fecundación hasta la forma adulta. Su propósito es describir y explicar las etapas de desarrollo de los metazoos, desde el estadio unicelular, hasta el estadio definitivo de diferenciación en que habiéndose formado totalmente los órganos del individuo, este podrá llevar una vida libre.
La embriología es una ciencia rica en cambios, empieza un desarrollo acelerado a partir de las ideas evolucionistas que surgen en el siglo pasado y con el uso de instrumentos ópticos adecuados, la idea de la concepción preformista es cambiada por conceptos evolucionistas (Lamarck, 1802 y Darwin, 1859)
El investigador alemán, Karl Von Baer fue el primero en hacer plausible la interpretación del desarrollo embriológico en 1828. Sus reglas de desarrollo embriológico describen una simple gradación de una semejanza generalizada en embriones de un amplio espectro de animales, muy singular antes que el embrión tome las características de las especies. Charles Darwin usó este concepto (Von Baer), señaló que de una similitud inicial divergen en una identidad individual, en el origen de las especies (1859). Para Darwin la similitud en embriones tempranos y su posterior divergencia constituyó evidencia de un ancestro común. Von Baer se opuso hasta su muerte a la interpretación de Darwin de su investigación, como evidencia de la evolución de la vida.
Ernest Hackel colaboró con Darwin en el uso de la embriología como argumento de evolución. Su Ley Biogenética hecha en 1866 fue traducida al inglés como "la ontogenia recapitula la filogenia", pero su interpretación correcta es una corta y rápida repetición de la filogenia.
Los biólogos modernos son cautos en sus explicaciones acerca del proceso de "recapitulación" y prefieren interpretar que las características embrionarias son retenidas y pasadas durante la evolución. Este cambio en la interpretación sugiere un retorno a la interpretación de Von Baer, que le ha sido agregada la idea del común ancestro promulgada por Darwin.
¿Por qué la cautela al hablar de recapitulación?
| La ontogenia contraria la filogenia, presenta un único genomio en su transcurso. El proceso filogenético puede y suele darse más de un genomio. | |
| En la ontogenia se da un concepto llamado telefinalismo, que implica un conocimiento del producto final del proceso. | |
| El registro fósil nos resulta, en ocasiones, insuficiente para poder inferir a partir de él, acerca de las estructuras blandas de organismos ya extintos. |
Se habla entonces de concordancia, donde el animal presenta, en diferentes etapas, una selección de características de sus posibles ancestros, la aparición de estas características estarían espaciadas de tal manera que concordarían con la aparición de las mismas en el tiempo geológico.
Las etapas de desarrollo
La ontogénesis o embriogénesis, de un individuo corresponde a una sucesión continua de cambios y variaciones mas o menos complejas. Su representación exacta exigiría una observación continua, hasta hace poco se pensó que era una tarea imposible, sin embargo, en la actualidad se ha estudiado minuciosamente el desarrollo embrionario del nematodo Caenorhbditis elegans a partir de una célula hasta el organismo completo de 959 células.
El desarrollo de la ingeniería genética es decir el arreglo dirigido de genes dentro de receptores biológicos, ha permitido un gran avance en la investigación del desarrollo embrionario en insectos, anfibios y mamíferos, llegándose a establecer el mecanismo por el cual las células logran una diferenciación en este proceso.
El desarrollo embrionario posee características que marcan fases durante las que se desarrollan los procesos del mismo orden. Estas etapas mas o menos virtuales, son de gran utilidad en la interpretación de los fenómenos.
El inicio del desarrollo ofrece aspectos semejantes en todas las clases de animales :
| Fecundación | |
| Segmentación | |
| Gastrulación | |
| Organogénesis |
La sucesión cronológica de estas fases y las transformaciones que sufren sugieren el origen común de todos los organismos animales.
Las ideas evolucionistas fueron el punto de partida para el desarrollo de la embriología descriptiva. Una vez conocido el curso del desarrollo normal, fue posible su estudio experimental y entonces nuevas preguntas necesitan ser respondidas:
¿"Por qué un cigoto es capaz de producir células, tejidos y órganos de estructura diferente si parten de un genomio?.
Dos teorías han dirigido el enfoque de las investigaciones:
- la teoría de la preformación
- la teoría de la epigénesis
La preformación, en el sentido estricto de la palabra significaba que el adulto se hallaba contenido por completo en el huevo. El desarrollo era sólo el engrandecimiento progresivo de ese adulto pequeño inicial. Como es de imaginar, que bajo esta forma, la teoría preformista no podría existir en la actualidad, surgiendo un cambio en estas ideas. Hoy esta hipótesis plantea que cada parte del futuro individuo no existe bajo su aspecto preformado definitivo, el huevo podría tener regiones definidas que posteriormente corresponderían a diferentes órganos del adulto. El huevo estaría formado por un "mosaico" de territorios o plasmas organoformadores cuyo destino sería fijado muy pronto.
Con estas ideas puede realizarse un experimento simple: al ser destruido un blástomero que correspondería al lado izquierdo, el blástomero que queda formaría un hemiembrión derecho, si los dos blastómeros siguieran su curso normal habrían producido un embrión completo, entonces el huevo se comporta de esta manera como un mosaico, Chabry (1887) y Coklin (1905) estudiaron huevos de Ascidias y descubrieron que las potencialidades totales son iguales a las potencialidades reales en los primeros blástomeros.
La epigénesis se inicia con Wolf (1759), médico alemán estudió el desarrollo embriológico del pollo y observó que el ave adulto no preexistía bajo una forma de miniatura en el huevo, sino que al contrario la estructura del embrión surgía poco a poco, mediante una sucesión de estadios, a partir de un estadio inicial muy simple. Actualmente la teoría de la epigénesis considera al huevo no como un mosaico de territorios ya fijados, sino como una "realización progresiva en la que cada etapa se halla condicionada por una etapa anterior más sencilla". En este caso las potencialidades totales del huevo son superiores a su potencialidad real. Esta hipótesis considera que la mitad del huevo podría dar un embrión completo, bien constituido aunque de menor tamaño y el huevo sería capaz de una regulación. En los embriones regulativos, parte del embrión puede quitarse y las células restantes pueden compensar la pérdida y dar como producto final un individuo completo.
Según estas dos teorías, ciertos huevos presentarían un desarrollo tipo mosaico y otros tipo regulación. Posteriormente los expertos demostraron que no hay antagonismo real entre ambas concepciones considerando como factor importante el tiempo.
Entonces tendremos que un huevo que presenta gran poder de regulación, pasado un tiempo limita sus potencialidades. Es decir un huevo relativamente homogéneo al principio capaz de regulación, mas tarde estaría formado por un conjunto de territorios bien determinados.
Como podemos pensar, la distinción entre embriones en mosaico y embriones regulativos ya no podía mantenerse y esta clasificación perdió su utilidad, pero tiene un significado histórico ya que dirigió la atención de los científicos de como se especifica el destino de las células en la embriogénesis. El fenómeno de la regulación centró la atención hacia el hecho de que grupos de células se comunican entre ellas y de este modo activan o desactivan programas específicos de expresión génica.
Polaridad.
El fenómeno de desarrollo en mosaico dirigió la atención en la localización de "determinantes" citoplasmáticos que actuarían como un mecanismo de especificación del destino de las células. Para el caso, el destino de una célula estaría determinado por información citoplasmática - determinante - que es heredada del huevo, lo contrario de una interacción inductiva: el destino de una célula queda especificado por información interna y no externa. En cualquiera de los dos casos debe producirse una diferencia inicial en la información del desarrollo, "una polaridad para el sistema". Esa diferencia inicial la podría dar una célula que le indica a otra lo que debe hacer o bien una región del citoplasma del huevo contiene información que no se encuentra en otro lado. Por lo tanto entender como se establece la polaridad en los primero estadios de desarrollo es muy importante para explicar como los ejes y esquemas de los tejidos hacen su aparición en la embriogénesis.
La estructura de un organismo esta controlada por sus genes. La información genética fluye del DNA al RNA y a las proteínas, durante el proceso de la transcripción, en el que la secuencia de bases existentes de una hebra del DNA cromosómico se transcribe, enzimáticamente, en una sola hebra de RNA. Después de la transcripción el RNAm pasa a los ribosomas, en donde actúan como matriz o patrón para la ordenación secuencial de los aminoácidos durante la síntesis de proteínas. En este proceso se da la réplica, transcripción y traslación de información, la réplica es la copia del DNA con formación de moléculas hijas idénticas, la transcripción es cuando el mensaje genético del DNA es transcrito en forma de RNA y llevado a los ribosomas y la translación es cuando el mensaje genético es descifrado y convertido en el alfabeto de 20 letras de la estructura proteica.
Al menos durante un siglo, e incluso mucho después que clarificara el papel que tiene el DNA en la herencia los mecanismos del control genético de estructura corporal permaneció en un complicado misterio. Durante los últimos años la mosca Drosophila ha proporcionado información al respecto, estos estudios han puesto de manifiesto la existencia de un tipo de genes de control del desarrollo, cuya función específica es la de determinar el patrón corporal. La combinación de la genética clásica y de la genética molecular esta demostrando como actúan estos genes. Se sabe que en Drosophila, no solo las estrategias generadas sino también los genes específicos que controlan el patrón corporal pueden tener equivalentes cercanos en vertebrados
El sistema de regulación genética consiste en tres clases de genes de control del patrón :
- En primer lugar , los productos de los genes de polaridad del huevo actúan definiendo las coordenadas espaciales del embrión, mediante la formación de gradientes de morfógeno en el huevo.
- A continuación los genes de segmentación permiten interpretar la información de posición proporcionada por el gradiente inicial de morfógeno. Estos genes dividen el embrión en una serie se segmentos - las unidades básicas modulares a partir de las cuales se construyen los insectos.
- Los productos de los genes de segmentación influencian la expresión de los genes homeóticos selectores, que mantienen la diferencia entre un segmento y otro.
Por medio de las actividades combinadas de los genes de segmentación y de los genes homeóticos selectores, a las células de cada segmento se les proporciona la información de valores posicionales, información que las células recuerdan y que guía su comportamiento posterior . Por último, en el interior de cada subdivisión segmental del cuerpo las células se comunican entre sí generando los detalles concretos de la estructura madura, gobernada aparentemente por la función de intercalación de la información.
Polaridad en Xenopus.
En el caso de Xenopus un huevo sin fertilizar presta un sólo eje de polaridad el eje animal - vegetal (A - V), sin embargo existen ejes adicionales de polaridad que se van sumando a medida que se va dando el proceso de desarrollo, la polaridad del eje A - V esta evidenciada por hecho de que las células del polo animal y vegetal tienen destinos diferentes. El polo animal origina la piel y el sistema nervioso, y el polo vegetal origina al sistema digestivo y la porción ecuatorial o zona marginal forma los tejidos mesodermales. Las moléculas responsables de la especificación de los destinos de las células animales ("determinantes") todavía se desconocen pero se ha demostrado que existen un sólo tipo especial de ARNms que están localizados a uno u otro polo del huevo. Uno de estos ARNms llamado Vg1, proviene de un gen que podría participar en la inducción mesodermal, ya que Vg1 esta localizado hacia el polo vegetal y codifica un factor de crecimiento. Inicialmente en el oogenesis, este ARNm se distribuye inicialmente en todo el citoplasma pero se transloca y se ancla en el cerca del polo vegetal durante el proceso.
Polaridad en Drosophila
Establecimiento del plan del cuerpo.
¿Cuál de los extremos en un embrión sería la cabeza ? ¿ Cuál su región caudal ? ¿ Qué determina estos ejes? ¿Cómo es la segmentación básica?
Durante muchos años estas preguntas permanecieron sin ser respondidas. A finales de la década de los 70 investigadores alemanes emprendieron investigaciones que continúan hasta hoy y han empezado a dar respuestas a estas interrogantes. Experimentos con mutaciones realizadas a través de la alimentación a moscas Drosophila machos adultas al ser apareadas su progenie mostró un amplio rango de defectos específicos que permitieron identificar tempranamente todos los genes y proteínas que controlan la ovogénesis en la mosca madre.
Los resultados revelaron que el eje corporal del embrión es actualmente determinado por moléculas suministradas por la mosca madre mucho antes de la fertilización. Existen células matriciales que ayudan al huevo al crecimiento dentro de la madre y estas producen también el movimiento de algunos de los componentes de ARN maternales hacia el extremo interior del huevo provocando una asimetría inicial. Parte de este ARN maternal proviene de un gen llamado "bicoide" que permanece atrapado cerca del punto de entrada marcando el extremo frontal (anterior) del embrión. Otro ARN maternal llamado nanos es translocado al otro extremo marcando la región posterior del futuro embrión.
Después de la fertilización el ARN maternal empieza a producir proteína bicoide, la cual difunde en el embrión en desarrollo y forma una gradiente de concentración. Al mismo tiempo en el otro extremo del embrión una proteína de otro gen maternal llamado "nanos" difunde dirigiéndose al centro. El nombre de estos genes son inspirados en su función - o carencia de ellos - dos mutaciones : un embrión mutante sin genes nanos carece de segmentación abdominal y también es pequeño, la palabra "nanos" deriva del griego que significa enano. Estas son gradientes opuestas y determinan el plan básico del cuerpo de la mosca. Cuando una mosca madre es incapaz de producir proteína bicoide produce embriones carentes de cabeza y tórax y los embriones presentan dos colas (bicaudales) así aparece el abreviado nombre bicoide. Sin embargo cuando el ARN bicoide es inyectado dentro del extremo frontal (anterior) de cada mutante estos se desarrollan normalmente y cuando los científicos introdujeron el mismo ARN en el extremo opuesto del embrión se desarrolló otra cabeza y otro tórax en el punto de inyección.
Genes de segmentación
¿Cómo exactamente estas gradientes proteicas ejercen su influencia?
Involucra una serie de genes que actúan en diversas zonas en el embrión :
Gradiente de concentración de proteína bicoide
|
Bicoide afecta a un gen : hunchback ARNm hunchback
(Un embrión sin genes hunchback carece de tórax y conduce a un forma jorobada).
|
ARN hunchback es distribuido por todo el embrión.
|
"Nanos" inhibe a ARNm hunchback (única función de nanos).
|
Sólo se sintetiza proteína hunchback en el extremo anterior (área controlada por bicoide).
|
La proteína hunchback se difunde al resto del embrión creando una segunda gradiente (responsable de la segmentación torácica y abdominal de la mosca)
|
Bicoide activa otros genes que controlan porciones mas pequeñas en el desarrollo del embrión en secciones progresivas.
|
3 Horas después de la fertilización el plan básico está trazado, han intervenido 30 genes en el patrón anterior - posterior y 14 en el patrón dorso - ventral.
|
Antes del término de la cascada (embrión con 14 bandas proteicas) se activan los genes homeóticos.
La expresión de estos genes se da luego de la expresión del cigoto pero antes de la diferenciación de cada célula en su segmento correspondiente.
Se desconoce si el modelo siempre es extensivo en su aplicación en otros animales. Los embriones tempranos de mosca son muy diferentes a los de mamíferos. Los embriones mosca son mas cercanos a una célula con múltiples núcleos suspendidos en su citoplasma, los embriones en mamíferos consisten de células separadas unas a otras por membranas, bicoide y otros morfogénos (sustancias difusibles que afectan la forma del cuerpo) pueden extenderse rápidamente en la masa fluida del embrión mosca, pero podrían tener largo tiempo de difusión a través de muchas membranas celulares en un embrión ratón o humano. Es probable que existan muchos "genes gradientes" involucrados en desarrollo de mamíferos que podrían trabajar en diferentes formas. No se ha encontrado muchos mecanismos trabajando en otros animales, se conoce un equivalente a bicoide en ranas llamado geosecoide, este descubrimiento podría indicar el proceso de determinación del patrón anterior (frontal) en embriones de rana.
Genes homeóticos y "homeoboxes"
Como hemos visto hasta aquí, durante las divisiones celulares que se dan después de la fertilización, las células toman diferentes formas, tamaños, funciones y potenciales, así como relaciones espaciales unas respecto a otras. Todos estos eventos se dan según factores definidos de tiempo. En algunas especies como el ratón las células que se forman en las primeras divisiones poseen todas el mismo potencial. A medida que la división celular procede, las células diferentes se orientan hacia la formación de solo un número limitado de tipos celulares adultos, por eso se dice son "determinadas". Las células determinadas están destinadas a diferenciarse en una forma específica y sus células hijas heredan este programa. La determinación viene seguida de una diferenciación, adquisición de diferentes fenotipos, ya sea a nivel morfológico o de propiedades bioquímicas. Al final es posible diferenciar más de 200 tipos celulares en diferentes en vertebrados. Hay evidencias como para indicar que existe un mecanismo de expresión génica diferencial, tanto en el tiempo como en el espacio, que determina el desarrollo y diferenciación, una de las metas al aproximarnos a este hecho a nivel molecular y de comprender la determinación y diferenciación es como de correlacionar los cambios espaciales y temporales con la expresión génica diferencial.
Genes homeóticos
Esta aproximación ha sido exitosa en Drosophila melanogaster, principalmente por que es posible la obtención de mutantes con defectos en estadios morfológicos tempranos. Los segmentos que se observan tanto en moscas en desarrollo como adultas se encuentran normalmente asociados con caracteres morfológicos específicos como el ojo, antenas, alas y patas. La genética clásica identificó un grupo de mutantes en Drosophila (antenapedia, bithorax, etc.), en los que los caracteres morfológicos particulares se desarrollan en posiciones anormales. En antenapedia por ejemplo aparecen patas donde deberían encontrase antenas. Los genes que están involucrados en este tipo de transformaciones morfológicas son denominados grupalmente como genes homeóticos, y las consecuencias de mutaciones homeóticas específicas están localizadas hacia uno o más segmentos. El descubrimiento de mutantes homeóticos permitió realizar ingeniosos experimentos que demuestran que el cuerpo de la mosca normal esta formado por un conjunto de regiones discretas, cada una de las cuales expresa un conjunto diferente de genes selectores homeóticos. Los productos de estos genes actúan como marcadores moleculares de dirección, equipando a las células con una especificación general de su valor posicional, por lo tanto, un mutante homeótico hace que un grupo completo de células estén desinformadas de su localización y por ello produzcan una estructura apropiada para otra región.
Los genes homeóticos controlan el desarrollo del sistema nervioso central así como también de estructuras musculares que se forman entre los segmentos. Estos genes no determinan el número de segmentos sino solamente el desarrollo morfológico.
Existen dos grandes regiones en el cromosoma 3 en Drosophila, cada una con 250 kb o más, que presentan los genes "antenapedia" o "bithorax". Dentro de estas regiones se encuentran lugares relacionados con la manifestación de dichas características. Algunos de estos lugares parecen regular la de las regiones en su totalidad, por ejemplo, la región ubx para los genes bithorax.
El funcionamiento de genes homeóticos particulares actúa en segmentos específicos al menos en parte de la transcripción específica. Los productos de estos genes son proteínas que poseen afinidad por determinadas secuencias de ADN y específicas en la regulación de otros genes. En conjunto estos experimentos demuestran que, así como la especificidad bioquímica el desarrollo morfológico esta controlado por un mecanismo de expresión génica diferencial.
También se han encontrado genes que se expresan en momentos específicos del desarrollo en organismos tan diversos como el maíz, nemátodes, erizos de mar, ranas y ratones. Se están utilizando bibliotecas de cDNA preparadas a partir de células que se encuentran en determinados estadios de desarrollo con el objeto de obtener clones que presenten RNAm en ese estadio particular. No obstante el potencial de obtener progreso rápido con la mayoría de los organismos no es alto, principalmente por que no es posible obtener información genética más rudimentaria. El nematodo Caenorhabditis elegans constituye una excepción ; no sólo porque se ha realizado un exhaustivo análisis genético, sino que ha sido posible determinar la línea que han seguido todas sus 959 células a lo largo del proceso de desarrollo a partir del cigote hasta el animal adulto. Entre los mamíferos el ratón es el animal más prometedor en cuanto a estos estudios, ya que existe abundante información y se conocen varias mutaciones que ocurren en el. Además es posible la obtención de células en estadios tempranos de desarrollo. En ratones y en algunos otros mamíferos, también es posible obtener líneas de desarrollo a partir de técnicas de cultivo a partir de embriones jóvenes.
Secuencias "homeobox"
Un gran número de genes conocidos que están involucrados en la regulación de la variedad de procesos en el desarrollo de casi todos los organismos eucariotas presentan una secuencia característica de 180 pb dentro de la región codificadora de la proteína correspondiente conocida como el "homeobox" por que fue identificada por primera vez en los genes homeóticos de Drosophila. La expresión de estos genes, que especifican la identidad de los segmentos que darán lugar a la cabeza, tórax y abdomen, se da luego de la expresión del cigote, pero antes de la diferenciación de cada célula en su segmento correspondiente . Desde que estas secuencias fueron descubiertas, han sido observadas en varios otros genes de Drosophila, en especial en aquellos genes que orientan a la células ha seguir determinadas en estadios tempranos del embrión. Virtualmente, cada uno de los genes presenta esta secuencia en Drosophila se expresa en un grupo particular de células embrionarias, y cada grupo de células presenta una combinación única de los productos de estos genes, algunos de estos genes parecen codificar factores reguladores de la transcripción que influyen en la expresión de otros grupos de genes "homeobox" así como de otras proteínas estructurales no reguladoras. De este modo el embrión se divide en un primer momento, gracias a la expresión de genes homeobox, en dos porciones : anterior y posterior , cada una con diferente potencial de desarrollo.
Donde quiera que se encuentre esta secuencia de 180 pb especifica una secuencia de 60 aminoácidos conocida como el homeodominio, actualmente considerado como un elemento que tiene alta afinidad por el ADN. Además de su localización casi invariable dentro de la proteína en la que se encuentran, es destacable resaltar su capacidad de formar estructuras helicoidales.
El análisis de varias secuencias de estas proteínas sugieren que existen similitudes y algunas diferencias entre vertebrados y Drosophila.
Existe suficiente evidencia como para asegurar que los homeodominios tienen una función importante en la unión de estas proteínas al ADN. La aparición de mutaciones puntuales en estas regiones disminuyen drásticamente el grado de afinidad de la proteína al ADN, mientras que alteraciones en otras regiones no afectan su afinidad. Una importante características de ciertas homeoproteínas es su habilidad de unirse a secuencias de ADN diferentes pero relacionadas, aunque con diferentes afinidades. Así esta especificidad diferencial permitiría que una homeoproteína regule la transcripción de uno o más genes.
Con respecto a la forma de unión de los homeodominios al ADN, se ha propuesto que la estructura helicoidal es crucial en la especificidad. Otras características de la proteína también podrían influir, pero en menor grado. En consecuencia, las propiedades reguladoras de la transcripción y/o unión al ADN están moduladas por pequeñas moléculas. Estas proteínas poseen regiones que las permite interactuar con otras proteínas de modo que podrían formarse complejos proteicos que actuarían como entidades funcionales. Una vez unidad al ADN, las homeoproteínas influyen en la transcripción desde sus correspondientes promotores blanco.
De acuerdo a los estudios realizados en los patrones de expresión de los genes homeóticos se puede ver al embrión de vertebrados como subdividido en campos anteroposteriores de células con diferentes capacidades de desarrollo, esta subdivisión del cuerpo embriónico precede a la formación de órganos o estructuras específicas. Aunque los homeodominios codificados por diferentes genes homeobox son muy similares entre si, las diferencias características en sus secuencias pueden ser utilizadas para identificarlos. Algunos homeodominios de mamíferos se asemejan fuertemente a aquellos producidos por genes de la mosca de la fruta.
Cuando los patrones de expresión de para muchos genes homeobox en embriones de ratón fueron analizados se realizaron importantes observaciones, se mostró que tanto en vertebrados como en invertebrados, los genes homeobox se agrupan en complejos o grupos en un cromosoma. En otras palabras. los genes homeobox están arreglados en un orden preciso, de izquierda a derecha, en la molécula lineal del ADN que forma un cromosoma. Además se descubrió que en ratón el orden de los genes homeobox en un grupo corresponde directamente a donde se expresa los genes. Los homeogenes localizados cerca del final izquierdo de un complejo son expresados en la parte posterior del cuerpo y los genes a la derecha son expresados mas cerca de la cabeza.
Todos los vertebrados tienen cuatro complejos homeobox , cada uno localizado en cromosomas separados. Estos complejos homeobox probablemente aparecieron durante la evolución a través de las duplicaciones de un grupo simple de genes homeobox en invertebrados. Consecuentemente cada humano por ejemplo, tiene cuatro genes que asemejan al gen Abdominal - B de Drosophila y otro que asemeja a Deformed.
Mutante Bithorax de Drosophila |
Espécimen nativo de Drosophila |
|
Mutante Antenapedia de Drosophila |
Drosophila nativa |


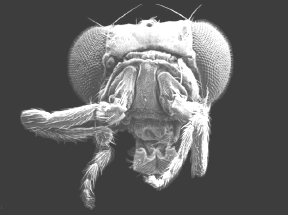
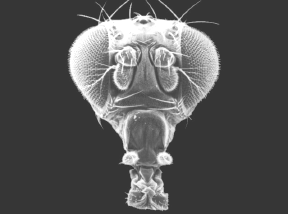
La secuencia homeobox está altamente conservada a lo largo de la evolución.
En 1990 los resultados de las investigaciones demuestran la existencia de este sistema de genes en embriones ratón y humanos. La notable homología de los genes que presentan una secuencia homeobox sugiere que han evolucionado por duplicación génica y divergencia. En el genoma de Drosophila muchos de estos genes se hallan agrupados muy estrechamente, lo cual también sugiere este origen por duplicación en tánden - por parejas - y divergencia. Por ejemplo, el complejo Antenapedia incluye bicoide, ftz, zerkült además del conjunto de genes homeóticos selectores. La agrupación de genes que presentan homeobox parece ser un reflejo de su historia y no de alguna necesidad fisiológica : las moscas que presentan reagrupaciones genéticas en las que el gen Ultrabithorax se separara de sus acompañantes del complejo Bithorax, no muestra anormalidades en su plan corporal.
Parece probable que a medida que fue evolucionando un plan corporal cada vez más complejo, se fueron añadiendo al genoma series de genes que contenían secuencias homeobox. Los fenotipos de los mutantes de Drosophila que carecen de genes determinados que presentan homeobox, proporcionan una visión de la posible estructura del organismo ancestral. Así eliminando los genes homeóticos selectores se produce un animal con numerosos segmentos idénticos, parecido a un ciempiés; especulativamente retrocediendo el curso de la evolución un paso más, al eliminar también los genes de polaridad de segmento y los genes de regla par, se puede alcanzar un hipotético ancestro, organizado como un nematodo, sin segmentación pero todavía presentando diferencias entre su cabeza y su cola. Por supuesto que un relato como este es altamente especulativo.
El descubrimiento de la homeobox ha iluminado algunas preguntas evolutivas importantes. Los genes que presentan homeobox se han encontrado no solo en insectos y otros artrópodos sino también el nematodo C. elegans, en anélidos (en sanguijuelas y en el gusano de tierra), en erizos de mar en cordados primitivos y en vertebrados, incluyendo ranas, pollos, ratones y en humanos. La secuencia homeobox está conservada con asombrosa exactitud a nivel proteico : por ejemplo una de las proteínas homeobox de Xenopus de la cual 59 de 60 aminoácidos son idénticos a la secuencia homeobox de la proteína antenapedia de Drosophila, a pesar que ambos organismos han evolucionado en forma independiente durante más de 500 millones de años. Esta conservación sugiere que pueden existir similitudes fundamentales entre los insectos y los vertebrados en cuanto al mecanismo que controla el plan corporal básico.
Los primeros Complejos homeobox deben haber aparecido muy temprano en la historia de la vida en platelmintos u otros organismos primitivos ancestros comunes tanto para el hombre como para los insectos. El alto grado de conservación de los complejos a lo largo de la evolución sugiere que una vez que una forma de especificar el eje anterior - posterior lo suficientemente eficiente apareció, fue más fácil producir nuevas formas corporales mediante la modificación de este sistema que desarrollar estrategias completamente nuevas.
Bibliografía.
- Basto Acosta Luís. (1992). Regulación genética del desarrollo y diferenciación del embrión. Separata para el curso de Histología y Embriología Animal. Departamento Académico de Biología. Universidad Nacional Agraria La Molina Lima.
- Brody Thomas (1999). Home Page : The interactive Fly © 1995. http://sdb.purdue.edu./fly/aimain/4matrnl.htm
- _____ (1998)Flies Invade Human Genetics Volume 12, No. 13The Scientist June 22,
- Córdova, J. H. (1990). Biología Evolutiva. Sollerita No. 8 Oct.- Dic.
- De Robertis, E. (1990). Homeobox Genes and the Vertebrate Body Plan. Scientific American. Julio 1990. Pag. 26 - 32.
- De Robertis (1992). Biología Celular. Cap. 16.
- Howard Hughes Medical Insitute. (1992). From egg to adult. Maryland USA.
- Mayr, E. 1988. Toward a New Phylosophy of Biology. The Bleknap Press of Harvard Uni. Press, England. 564 pgs.
- Nüsselein - Volhard Christiane. (1996). Scientific American. August 1996. Pag. 38 - 43.
- Swan, L. W. (1990). The Concordance of Ontogeny with Phylogeny. BioScience vol. 40 No 5
- Encyclopedia of Life Sciences http://www.els.net
Drosophila Web Sites
The Interactive Fly http://sdb.bio.purdue.edu/fly/aimain/1aahome.htm
Flybase (data) http://flybase.bio.indiana.edu
Drosophila virtual library (Internet resource list) http://ceolas.org/fly/
Flybrain (atlas and database of nervous system) http://flybrain.neurobio.arizona.edu/
FlyView (image database) http://pbio07.uni-muenster.de/Home.html
Berkeley Drosophila Genome Project (encyclopedia of sequence data) http://fruitfly.berkeley.edu/
DRES (Drosophila-related expressed sequences) http://www.tigem.it/dros.html