VOLVER A PROGRAMA
VOLVER A PROGRAMA
VOLVER A PROGRAMA
VOLVER A PROGRAMA
VOLVER A PROGRAMA
VOLVER A PROGRAMA
VOLVER A PROGRAMA
VOLVER A PROGRAMA
VOLVER A PROGRAMA
VOLVER A PROGRAMA
VOLVER A PROGRAMA
VOLVER A PROGRAMA
VOLVER A PROGRAMA
VOLVER A PROGRAMA
VOLVER A PROGRAMA
VOLVER A PROGRAMA
VOLVER A PROGRAMA
VOLVER A PROGRAMA
VOLVER A PROGRAMA
VOLVER A PROGRAMA
VOLVER A PROGRAMA
VOLVER A PROGRAMA
VOLVER A PROGRAMA
VOLVER A PROGRAMA
VOLVER A PROGRAMA
VOLVER A PROGRAMA
VOLVER A PROGRAMA
VOLVER A PROGRAMA
VOLVER A PROGRAMA
VOLVER A PROGRAMA
VOLVER A PROGRAMA
VOLVER A PROGRAMA
VOLVER A PROGRAMA
VOLVER A PROGRAMA
VOLVER A PROGRAMA
VOLVER A PROGRAMA
VOLVER A PROGRAMA
VOLVER A PROGRAMA
VOLVER A PROGRAMA
VOLVER A PROGRAMA
VOLVER A PROGRAMA
VOLVER A PROGRAMA

UNIVERSIDAD NACIONAL AGRARIA LA MOLINA LIMA-PERU
Departamento de Biología
Curso CC6012
"Principios de Evolución"
Profesor Luis Basto Acosta
PROGRAMA
| 1.-INTRODUCCION | 17.-SE AGREGARAN NUEVOS CAPITULOS |
6.-MONOMEROS |
|
9.-PROTOBIONTE |
|
| 11.-LA REVOLUCION DARWINIANA | |
| 12.-CRONOLOGIA DE LOS SIETE MOMENTOS ESTELARES DE LA EVOLUCION | |
| 13.-LAS EXTINCIONES | |
| 14.-BIOLOGIA COMPARADA: HOMOLOGIA Y ANALOGIA | |
| 15.-EL ARBOL DE LA VIDA: FILOGENIA | |
| 16.-EVOLUCION HUMANA |
![]()
![]()
1.-Introducción |
Nada tiene sentido en la biología si no es bajo el prisma de la Evolución.
Theodosius Dobzhansky
Pocas ideas han cambiado tan profundamente nuestra visión de la naturaleza como la misma idea de cambio que está implícita en la evolución de los seres vivos. Los organismos se agrupan en unidades naturales de reproducción que denominamos especies. Las especies que ahora pueblan la Tierra proceden de otras especies distintas que existieron en el pasado, a través de un proceso de descendencia con modificación. La evolución biológica es el proceso histórico de transformación de unas especies en otras especies descendientes, e incluye la extinción de la gran mayoría de las especies que han existido. Una de las ideas más románticas contenidas en la evolución de la vida es que dos organismos vivos cualesquiera, por diferentes que sean, comparten un antecesor común en algún momento del pasado. Nosotros y cualquier chimpancé actual compartimos un antepasado hace algo así como 5 millones ańos. También tenemos un antecesor común con cualquiera de las bacterias hoy existentes, aunque el tiempo a este antecesor se remonte en este caso a más de 3000 millones de ańos. La idea de evolución por modificación y derivación de nuevas especies implica la existencia de antepasados comunes para cualquier par de especies. Hay un antepasado común del hombre y el chimpancé, y del hombre y las bacterias. La evolución es el gran principio unificador de la Biología, sin ella no es posible entender ni las propiedades distintivas de los organismos, sus adaptaciones; ni las relaciones de mayor o menor proximidad que existen entre las distintas especies. La teoría evolutiva se relaciona con el resto de la biología de forma análoga a como el estudio de la historia se relaciona con las ciencias sociales. La famosa frase del genético evolucionista Theodosius Dobzhansky que abre este tema, no es más que una aplicación particular del principio más general que afirma que nada puede entenderse sin una perspectiva histórica.
![]()
![]()
2.-Historia del pensamiento evolucionista
żCuáles fueron los conocimientos iniciales sobre la evolución biológica?
Los temas del origen de las especies y el de la evolución del hombre fueron considerados tabúes o "malditos" hasta hace unas pocas centenas de ańos. Se pensaba que eran irresolubles y no accesibles a la ciencia. Y es que era casi imposible lidiar con dogmas de base animista, enraizados con el hombre primitivo desde siempre. Enraizamiento comprensible si se tiene en cuenta la existencia de hechos que afectaron profundamente al emergente Homo sapiens: como la acaecencia de la muerte, la procreación, y los devastadores eventos naturales que perturbaban su bienestar físico. Su consternación aumentaba porque no entendía las leyes de estos sucesos. La necesidad de un esclarecimiento, lo indujo a pensar que eran producidos por alguna Voluntad Divina. Y como quizás era mejor mantener una relación amistosa con este ente superior, desarrolló rituales, prácticas amistosas y consideraciones animistas finalistas, de ancha base que explicaban todo, incluyendo la creación del hombre y las especies (James, 1981: 40-41).
Así se continuó por muchos ańos. Pero con el andar del tiempo algunas religiones tuvieron que argumentar más. Santo Tomás de Aquino (1225 - 1274 ańos D.C.), en Summa Theologiae, logró fundir las ideas aristotélicas con las evolutivas de base platónica, adecuadamente amparadas en el equilibrio esperanzador que proporciona la fe animista. Su punto de vista era concordante con el hecho de que el hombre y las especies habían sido creadas en forma independiente, aunque bajo formas perfectamente elaboradas (Cid et al., 1965:638).
 |
CHARLES DARWIN "El origen de las especies" |
La consolidación paulatina de la ciencia (búsqueda de la verdad por métodos exactos y razonados) como construcción mental de las emergentes civilizaciones iba, sin embargo, imponiendo poco a poco, exigencias impostergables. Charles Darwin encaramado sobre los hombros de notables pensadores precedentes, como Aristóteles (384-322 A. C., que creía en la especiación, por generación espontánea), Mapertuis (1698-1759, que creía que la especiación se producía por la acumulación de anomalías fortuitas), Lamarck (1744-1829, que creía en la herencia de los caracteres o rasgos adquiridos y que las especies se transformaban por el uso y el desuso), Laplace (1749-1827, estudioso de la evolución de planetas y estrellas) y otros; logra publicar en 1859, el Origen de las Especies, donde postulaba la evolución de las mismas a partir de un antiguo ancestro común. En esta obra seńalaba además que la variabilidad de las mismas, hecho necesario para adaptarse a distintos nichos ecológicos podía ser explicada por el trabajo modelador de la selección natural. Estas ideas se originaron a partir de las observaciones obtenidas de su trabajo como naturalista, en un largo viaje realizado entre Australia y América del Sur, pasando por Africa, entre 1831-1836. El Origen de las Especies al recibir la aceptación rápida y unánime de la comunidad científica (no sin cierta resistencia de parte de los credos animistas), supuso a la vez, una reorientación filosófica y biológica sobre el origen y el destino del hombre.
Esta teoría sin embargo, tenía algunos defectos, ya que Darwin suponía que todos los cambios evolutivos ocurrían por la selección natural de pequeńas diferencias individuales que aparecían generación tras generación, en cualquier especie. Y aunque pequeńas, dándoseles el tiempo suficiente, podían dar lugar a grandes cambios evolutivos. Sin embargo, los saltos especiativos existen (equilibrio punteado); y parecen ser dependientes de circunstancias ambientales específicas, promotoras de una acción redoblada de las fuerzas de la selección natural (Cronin et al., 1981:119-122).
Darwin al defender lo suyo afirmaba, que la ausencia de fósiles que confirmaran eslabones intermedios se debía a:
- un registro fósil imperfecto (muchas formas intermedias "no encontradas," se hallarían en los sedimentos de los fondos marinos)
- sólo ciertas formas orgánicas son posibles de ser preservadas
- sólo las especies dominantes y ampliamente distribuidas son recuperadas frecuentemente, y éstas por ser halladas en lugares distantes dan la impresión de haber sido creadas de súbito, y
- dentro de las múltiples gradaciones intermedias, unas pocas especies habrían sufrido la presión de los cambios evolutivos (Darwin, 1936 [1859]:350-357).
żQué pensaba Teilhard de Chardin?
En los ańos siguientes se descubrieron cráneos de Homo erectus arcaicos en Düsseldorf (1856), Java (1891), Pekín (1920), y Sudáfrica donde Dart en 1924, encontró los restos fósiles del primer australopitecino. Abundaban ya también a ojos vista, restos fósiles de especies distintas al género Homo, correspondientes al período Cámbrico, que ayudaron a esclarecer los nexos evolutivos existentes entre ellas. A estos eventos, se sumarían conocimientos más elaborados relacionados con la física einsteniana, la génesis del planeta Tierra, la tectónica de placas, la deriva continental, y se empezaron a relacionar las glaciaciones con las grandes migraciones humanas. Estos hallazgos al plantear mayores exigencias, instaron a los predios animistas a responder con mayor altura. La Teología alcanzaría, bajo esta presión, altas cimas en el pensamiento de Teilhard de Chardin. Teilhard, que según el mismo, era más naturalista y filósofo, que físico, imprimió una nueva orientación al discurrir animista: aceptó la evolución del hombre y las especies aunque de un modo peculiar; es decir siguiendo la dirección de la ortogénesis (divergencias evolutivas diferentes para el hombre, las plantas y los animales) las cuales coalescerían, al final en un todo, de un modo determinista y finalista (el punto Omega), en una unión que sería más psíquica que biológica (Teilhard de Chardin, 1974: 15-85). Los planteamientos de este rara avis religioso, no lo salvarían del exilio y la marginación aplicada a él, por sus superiores, por el solo hecho de pensar en forma diferente (Combes, 1975:171-177). Por si fuera poco, Teilhard ha recibido en los últimos ańos acres críticas de connotados evolucionistas que le increpan su falta de racionalidad. Paradójicamente James Lovelock parece haber vestido de frac los conceptos de biosfera (capa viviente que circunda la tierra), de Teilhard. Lovelock coincide en que la Tierra en su conjunto, es un organismo homeostático viviente y ha aportado que esta superestructura puede ser estudiada científicamente si se considera la existencia de moléculas universales que recorren el mundo como la clorofila, la glutamina sintetasa y otros. Aparte de que existen organismos universales que fijan el nitrógeno, reducen los sulfatos, y realizan funciones de respiración y de fotosíntesis. Además recientemente ha quedado establecida la existencia natural, a amplia escala de la transferencia y/o intercambio de partes de genes o genes completos entre especies: transferencia genética horizontal, incluyendo algunas remotamente relacionadas (Miller, 1998:47), sugiriendo que por lo menos en el ámbito molecular y a despecho de las diferencias inter-específicas existe la tendencia a formar un todo unificado. Este mecanismo apoya las hipótesis ortogenéticas (Syvanen, 1994:256). Con lo que se puede afirmar que el buen Teilhard goza actualmente de buena salud
żCuales fueron los aportes de la genética a la Teoría Sintética de la evolución biológica?
 |
GREGORIO MENDEL |
Los conceptos sobre las infinitas gradaciones de las especies se aclararían con el conocimiento de la existencia de unidades genéticas alternantes (alelos) inductores de variadísimas formas, que siguen las leyes de la segregación y la transmisión independiente, descubiertas por Mendel en 1865. Es decir la posibilidad de escoger lo más conveniente que existe. Y si un nicho ecológico impone una presión selectiva persistente, es posible tener ciertos tipos morfológicos dominantes; y no necesariamente, toda la especie ( Ayala, 1980:21-57).
Los aportes de Mendel necesitaban ser integrados, lo que fue realizado por Gaylord Simpson en 1944, cuando publicó Tempo and mode in evolution: Tempo (frecuencia evolutiva o cantidad de cambio morfológico con relación a un standard: inercia vs. evolución lenta o rápida) y Mode (patrón de la evolución: gradualismo filogenético vs. "equilibrio punteado") . En esta obra se integraban conocimientos de paleontología, genética, selección natural y matemáticas. Quedaban en ella unidas, los aportes de Dobzhansky (versión integradora de genética y selección natural, accesible al público), los de Fisher, Haldane, Chetnerikov ( teóricos biométricos que definieron en términos matemáticos las frecuencias evolutivas), las de, de Vries (opositor de la selección natural), las de Pearson y Weldon (biometristas), las de Stebbins (botánica), las de J. S. Huxley (zoología), y las de Mayr (sistemática).
La síntesis o Teoría Sintética exigió a Simpson definir a los moduladores de la frecuencia evolutiva (determinantes genéticos) como son:
- Tamańo poblacional: pequeńo y grande
- Frecuencia de mutaciones
- Variabilidad: alta y baja
- Selección natural y variantes: selección sexual, selección direccional etc.
- Efectos de las mutaciones
- Tamańo de las generaciones y otros (Simpson, 1944).
Los gaps fósiles quedaban ahora así explicados.
Hoy se conoce además, que la frecuencia evolutiva de la mayoría de las células procariotas es baja, con relación a las eucariotas. Del mismo modo, la frecuencia evolutiva de los mamíferos, por ejemplo es 5 veces más alta que la de otros animales. En general la velocidad de las frecuencias evolutivas guarda una relación directa con la mayor complejidad de los organismos (Schopf, 1944: 6735-6742).
La genética abrió nuevos campos. Múltiples trabajos realizados en bacterias y en Drosophila melanogaster, interpretados adecuadamente por científicos de renombre como Monod, Jacob y Dobzhansky, han colocado al azar en el rango de dirimente en muchas alteraciones moleculares (mutaciones) (Monod, 1971:133-136, Dobzhasky, 1979:6-16), como también en ciertas etapas evolutivas de los organismos pluricelulares, signadas por fortuitas extinciones, que eliminan masivamente a las especies mejor dotadas (Raup, 1944:6758-6763). Así, el Homo sapiens no sería el producto de un determinismo finalista, sino de la síntesis del azar y la selección natural
żQué aportes tuvieron la física, la química y otras ciencias en la comprensión de la evolución?
La materia y la energía también ofrecen instructivas lecciones evolutivas. Un conocimiento más prolijo de las dataciones geocronológicas de fósiles, fue proporcionado por la física, al conocerse la fusión y la fisión nuclear. Los conceptos sobre fisiones nucleares espontáneas y radiaciones, han permitido en la actualidad, el uso de isótopos de gases raros (químicamente inertes), como el Argón y el Xenón, que ofrecen fechas más precisas. El tiempo geológico se mide por la velocidad de la conversión de un isótopo en otro (Allégre y Schneider, 1994:36-45). La física también ha permitido explicar en parte la quilaridad de los seres vivientes dentro del contexto del principio de incertidumbre de la física cuántica (Hegstrom and Kondepudi, 1990:58-67). Del mismo modo, esta rama de la ciencia, ha proporcionado la idea de que podríamos ser sólo organismos circunstanciales, probabilísticos en esencia, posibles de habitar el universo sólo en ciertas áreas del espacio-tiempo (en los periódos expansivos del ciclo del universo) (Dicus et al., 1983:74-85). Según Miró-Quesada la teoría de la expansión del Universo tiene en la existencia de la radiación de fondo, que recorre el cosmos en toda su extensión, una prueba concreta (Miro-Quesada, 1991:441). No dejan de ser importantes los conocimientos recientes sobre la evolución de los elementos, y la formación de compuestos carbonados a escala interestelar. Los meteoritos trayéndonos materia orgánica consolidada de cometas y estrellas lejanas (Cronin and Pizarello, 1997:951-954), han sugerido la posibilidad de emergencia de vida en áreas físicas distintas al del planeta Tierra.
El avance de los conocimientos en torno a la evolución ha sido vertiginoso en los últimos ańos y su espectro abarca hoy en día el mundo inorgánico, los reinos animal y vegetal, la conducta, el lenguaje, la cultura etc. El resultado de todo esto ha sido desde el punto de vista onto y filogenético de las especies, de una gran riqueza conceptual. Más integraciones son necesarias, sin embargo. Könberg en 1987, enfatizó la necesidad de integrar la Bioquímica con otras ciencias, y de modo especial con la Biología (Körnberg, 1987: 6888-6891). Diez ańos después los frutos de esta recomendación son reconfortantes, pero como lo reitera Castro de la Mata, los logros alcanzados deben ser siempre interpretados con cautela, porque los detalles factuales, van cambiando algunos aspectos, pero lo que en general se conoce, es poco. Los conocimientos que en la actualidad se tienen del hombre deben ser interpretados dentro del contexto de que él, sólo constituye una pequeńa fracción de los seres vivos (Castro de la Mata, 1987:5-8).
Mucho de lo precedente ayudó a Oparin y a Haldane en 1920, a plantear la posibilidad de la emergencia de la vida a partir de condiciones prebióticas, en atmósferas reductoras que consistían de mezclas de CO2, N, CH4 y NH3. Estas ideas fueron recogidas por Miller, quien en 1953, logró obtener en forma artificial aminoácidos y diversos compuestos orgánicos básicos para la conformación de la vida (Oparin, 1979:12-89, Miller, 1953:528). Por esta misma época, Schopf y otros hallaron células procariotas fósiles de 3,500 Ma, de existencia (Schopf, 1979:52-55). De aquí, el enlace evolutivo de las especies vivientes a un organismo común ancestral es cercano. Aunque en el borde de la cercanía, los fósiles ya no existen.
Donde no existen fósiles concretos, emergen conceptos abstractos como el de microfósiles (cualquier molécula cuya estructura o función contemporánea permita o proporcione una guía de su historia evolutiva), lo que ha permitido establecer marcos conceptuales como el del "mundo del RNA". La importancia de estas ideas, es la de ser guías en la elaboración de los experimentos que a escala molecular se vienen realizando en los últimos ańos. Experimentos que a su vez, han tenido un mayor impulso con los hallazgos de la "revolución woesiana", que examinó las relaciones de oligonucleótidos hallados y secuenciados en los ribosomas de procariotas ancestrales; lo que ha permitido establecer relaciones sólidas en la secuencia evolutiva de plástidos en vegetales, y de mitocondrias en animales (Woese and Fox, 1977: 5088-5090). Del mismo modo ha permitido establecer una relación celular ancestral, la más antigua que se conoce; planteando el concepto que el sistema primitivo genético ancestral (cenancestor o progenote) no habría sido muy diferente al de las células actuales, es decir que las diferencias entre procariotas, eubacterias y eucariotas, sólo serían entendidas dentro del concepto de que a partir del progenote se habrían dado divergencias evolutivas en razón del hábitat particular de cada una de ellas (Doolittle and Brown, 1994:6721-6728).
Como en todo tiempo y lugar se cuecen habas, la identificación de las arqueobacterias por métodos no convencionales le costó a Carl Woese, el ostracismo y la pérdida de su reputación por 15 largos ańos. La propia ciencia no aceptaba otro método que los cultivos, para la identificación plena de las bacterias (Morell, 1997:700-702).
Fueron importantes también, por esta época, los finos conceptos vertidos por el físico Schrödinger, sobre temas biológicos. Este científico tipificó a los organismos vivos como manipuladores y acopiadores de energía teniendo como base una suerte de mecanismo entrópico negativo, generador (en contraposición al desorden entrópico que propugna la segunda ley de la termodinámica), de códigos de orden sumamente complejos, leáse: leyes que rigen la formación de códigos genéticos, leáse leyes que rigen la formación de los elementos, y cuyos cimientos deben residir en alguna desconocida ley universal que gobierna bajo un solo puńo los fenómenos biológicos, químicos y físicos en todo el Universo. No menos notable fue su predicción 15 ańos antes de los descubrimientos de Watson y Crick, en el sentido que los genes eran capaces de preservar sus estructuras, porque el cromosoma que portaba toda la información se comportaba como un cristal aperiódico. Estos cristales estarían compuestos de una sucesión de pequeńos elementos isoméricos, la naturaleza de los cuales constituiría el código hereditario. Vastas posibilidades combinatorias son posibles, si los mismos dispusieran del uso de solo 2 símbolos similares al código Morse ( Schrödinger, 1956:22-75)
żY el origen del hombre?
El paso de procariotas a eucariotas, necesitó más de 2,000 Ma. Las procariotas sin embargo no han desaparecido, y su vigencia y persistencia a la fecha, es sinónimo de vigor. En este aspecto, los insectos emergidos hace 350 Ma, no se quedan atrás con sus 2x106 especies. El Homo sapiens sin embargo en apenas 200,000 ańos ha ańadido a estas diversificaciones un nuevo concepto, el del progreso (cambio de un todo, o una característica precedente, a otra mejor, y no necesariamente siguiendo líneas direccionales, graduales y uniformes).
En el presente resumen histórico progreso y evolución se toman como sinónimos. Evolución implica eficacia biológica, complejidad y abundancia (Dobzhansky, 1969: 180-205). En su sentido más amplio evolución significa también cambio, sucesión y transformación de todas las cosas en el tiempo, conducentes en la mayoría de los casos a formas más avanzadas y complejas (Silva Santisteban, 1988:35). Un buen ejemplo de evolución o progreso, lo constituye el desarrollo del lenguaje y el cerebro, mediadores de la cultura moderna, con las que el hombre ha pasado a dirigir su propia evolución.
De la ciencia macro (Antropología física), que identificaba con facilidad esqueletos casi completos, se ha ido pasando poco a poco (ante la exigencia de hurgar más y más, en el pasado), a la interpretación de la estructura y función de piezas fósiles cada vez más pequeńas. Los especialistas se dan ahora el lujo de elaborar interpretaciones morfológico - funcionales a partir de minúsculos segmentos de especies vegetales, o de exiguos fragmentos óseos: fósiles de homínidos y seres humanos. Y, en la medida en que los descubrimientos objetivos son conocidos por el gran público, existe una coincidencia cada vez más apretada entre lo que se predijo, y lo que se descubre. En este sentido, la interpretación funcional de los restos fósiles de Ardipithecus ramidus (White et al., 1994:306-313), de Australopithecus anamensis, y otros constituyen por su exquisitez y precisión, un auténtico state of the art. En este aspecto Darwin no se quedaba atrás. Con pocos elementos a la mano, en el Origen de las Especies predijo los cambios que ocurrirían en caninos y molares en ancestros del género Homo. 100 ańos después, al identificarse las arcadas dentarias de australopitecinos y homínidos, los hechos le dieron la razón (Darwin, 1936[1859]: 345-357).
La necesidad de ir más atrás, nos ha llevado a la consolidación de la genética molecular. El secuenciamiento del mtDNA (ácido desoxiribonucleico mitocondrial) de un fragmento óseo de un Homo neanderthalis, ha permitido casi establecer que era una especie diferente (Krings and Pääbo, 1997:19-30), sugiriendo la existencia de múltiples planes formadores de homos, varios de los cuales quedaron en zonas sin progreso, en distintas áreas geográficas del mundo, reafirmando el rol del azar y la selección natural al determinar la preeminencia de ciertas especies en ciertos períodos temporales.
Mucho se trabaja hoy en el desentrańamiento del misterio del diseńo de las formas corporales ancestrales. El descubrimiento del complejo de genes Hox / HOM regulando la secuencia céfalo-caudal de la mosca de la fruta, es en este sentido un pequeńo gran paso hacia delante (Mcginnis y Kuziora, 1994:22-28). Del mismo modo se trabaja febrilmente también, en el estudio genético de las fases evolutivas de la especie Danio rerio ("zebrafish"), un pez multicoloreado de 2,5 cm que habita en las aguas del río Ganges. Los huevos de esta especie son translúcidos, y se piensa que el seguimiento evolutivo de sus embriones hasta constituir larvas capaces de nadar, e igualmente translúcidas (lo que se logra en 5 días), permitirá conocer la secuencia de la génesis de órganos internos y estructuras óseas, derivándose de una sola célula (Postletwait and Talbot, 1997:183).
Así, contemplando reflexivamente la evolución del Universo y sus diversos constituyentes desde una butaca edificada sobre más de 300 ańos de conocimientos científicos, el tema de la evolución aparece firmemente consolidado en sus líneas generales, pero casi acabado al mismo tiempo, porque muchos hechos futuros, se pueden atisbar como nunca antes y hasta se pueden predecir.Lo que recién empieza a nacer entonces, es la perspectiva de la evolución.
Por lo pronto se ha propuesto un nuevo tipo de ética basada en la conservación y en la promoción de la eficacia biológica de la especie Homo sapiens sapiens. Dentro de este nuevo código o cuerpo de leyes biológico serían incorporadas ideas o principios cualesquiera sea su origen, siempre y cuando resistan los test, a que serán sometidos por la ciencia (Monod, 1971:183-188).Porque hasta la emergencia del Homo erectus, hace 2 Ma, las fuerzas ciegas, salvajes y anéticas de la selección natural habían obrado sin tregua y sin pausa, modelando y seleccionando a las especies que estaban destinadas a sobrevivir, y eso en cierta forma era el sentido de la evolución. Hoy se considera la posibilidad de que las supuestas formas finales morfológicas (Homo sapiens sapiens, por ejemplo) sean circunstanciales; es decir que estarían determinadas por el azar y la necesidad y no por algún plan maestro intencionado (Jacob,1986:286-297). De ahí en adelante se ha continuado avanzando por el estrecho sendero que dejaba la fuerte pugna establecida entre el ambiente y la cultura, y conforme han ido pasando los ańos un claro vencedor ha emergido: el del claro convencimiento de que la evolución será dirigida mayoritariamente por concepciones culturales (Dobzhansky, 1974:15-23).
ż Cómo enfrentar el futuro del hombre con la nueva biotecnología?
Y esto implica una gran responsabilidad en el momento de la toma de decisiones, porque los programas eugenésicos a gran escala por ejemplo, factibles de ser implantados por determinados gobiernos generarán fricciones, las sociedades estarán a favor y en contra, cada punto de vista será el mejor. Las decisiones tomadas en estas circunstancias, bajo la influencia de determinados patrones culturales podrían al lado del logro de efectos benéficos de corto alcance; a inducir dańos genéticos irreversibles en parte, o en la totalidad de la humanidad. La perspectiva de la evolución por estas razones es nueva, y de profundo significado en los ámbitos filosófico, biológico y ético.
Casi parece pues que ha terminado un ciclo. El nuevo empieza con la necesidad inmediata de alargar la duración de la vida. A largo plazo emerge la posibilidad de crear vida, en forma planificada a partir de materia orgánica y / o inorgánica, y no necesariamente con las formas que ya conocemos. Obviamente se espera que los descubrimientos científicos de los próximos ańos tengan que ver con los tendientes a alargar el cronos del genoma. El balbuceo inicial en este sentido esta dado por la elaboración de mapas completos de genomas de vertebrados y otras especies. Según Bernardi, el análisis de los secuenciamientos iniciales de nucleótidos, esta revelando grandes divergencias entre isocoros de plantas y animales, ofreciendo este modo las líneas básicas para los estudios diferenciales de la longevidad (Bernardi, 1995: 445-476). Del mismo modo son importantes los conocimientos que relacionan la longevidad y ciertas funciones de los telómeros (extremo distal, especializado y renovable de causa cáncer y envejecimiento (Zaklan, 1995: 1601-1606, Greider, 1996:337-365). De otro lado la elongación artificial de los telómeros, prolonga la juventud de las células y permite un mayor número de divisiones celulares (Bodnar et al., 1998:349-352). A éstas, habrá que ańadirles lo que se pueda obtener de la clonación humana, (Wilmut, et al,. 1997:810-813), especificada tal vez en la creación de autoórganos (corazón, rińón, pulmones etc.), que servirán para el reciclaje temporal de los dańados. Aunque probablemente este esquema sea abandonado, porque sólo tal vez llegue a duplicar la edad promedio actual.
La superación de esta etapa podría promover la gestación de cambios conceptuales promovidos por el normal avance de la ciencia, pero también por el claroscuro de la mente humana, la ambición y el poder. Y, tal vez sin importar los medios, arribaremos a edades más amplias combinando lo anterior con la creación de verdaderos cyborgs (humanos con reemplazos de ciertas áreas corporales, por mixturas de prótesis metálicas sumamente sofisticadas, y autoórganos obtenidos por clonación).
Esto pese a todo será insuficiente, y entonces se comprenderá que el verdadero alargamiento de la vida tendrá que ver con la adquisición por parte del soma humano de formas de energía diferentes, quizás de duración ilimitada, lo que exigirá de hecho que se tenga que abandonar el concepto de que el Homo sapiens tenga necesariamente la conformación corporal que ahora detenta. Tal vez el soma humano sea reemplazado totalmente. Estas conformaciones corporales serán muchas y mayormente serán dictadas por la adecuación a ambientes particulares y específicos (Norton et al., 1986: 190). El alcance de la imaginación y la cultura fijarán la otra cuota.
Todo cuanto se haga, se descubra y / o se sintetice de hoy en adelante tendrá este fin. Lo que se presenta a continuación es eso, hechos mayoritariamente, con algunos eventos hipotéticos, conformando sólo la parte estructural, mecánica de la evolución, pero no el fin último, ni las interpretaciones finales. No hay hechos finales, solo existen interpretaciones y perspectivas (Castro de la Mata, 1987: 5-8).
![]()
![]()
3.-Evolución del universo
 |
COMETAS EN FORMACION |
El estado actual de conocimientos en torno a la génesis y evolución del Universo en el que vivimos, está basado en una interpretación teórica de consenso (mayormente de la ciencia física), de algunos fenómenos concretos que involucran la materia y la energía.
Estos fenómenos han gestado teorías generales, las mismas que se han ido perfeccionando a velocidad creciente, a partir del siglo XVII. El proceso de depuración de las mismas ha devenido en la preeminencia de dos de ellas, siendo la tendencia de éstas a fusionarse (Hawking, 1992: 201-230).
La primera de ellas se nutre de las leyes de la física clásica a gran escala, es decir de la que lidia con leyes que regulan el movimiento de los planetas, los campos gravitatorios, las velocidades a gran escala, etc. Este tipo de física, con Newton como mentor principal, es muy exquisito en sus exigencias, de hechos rigurosamente exactos en su comprobación. Albert Einstein, creador de la teoría de la Relatividad General y Especial, se adhería a esta rigurosidad, al hacer notar siempre que "Dios no juega a los dados" (Einstein, 1980:47-74).
La segunda teoría por el contrario está más involucrada con la física de escala microscópica, la que gobierna las leyes del movimiento, la energía y la materia de las partículas elementales (de las que se cree que ya no pueden ser divididas), que constituyen los núcleos atómicos: la física cuántica. Este tipo de física estudia 4 fuerzas:
- La fuerza gravitatoria: fuerza de atracción ejercida por las masas y la energía
- La fuerza electromagnética, que interactúa con partículas cargadas eléctricamente como quarks y electrones
- La fuerza débil: responsable de la radioactividad emitida por los procesos de fisión nuclear, y que actúa sobre partículas elementales de espin ˝ ,como fotones y gravitones y
- La interacción de la fuerza fuerte: que mantiene férreamente unidos dentro del núcleo, a fotones y neutrones (Hawking, 1992:101-112).
La física cuántica a diferencia de la clásica, no sólo trata de sucesos rigurosamente exactos y determínisticos, sino tambien de hechos probabilísticos que acaecen en complejos del tipo . electrón-antielectrón, quarks- antiquarks (Harari, 1983:48-60). Las antipartículas, habitúes de la antimateria, se tornan verificables durante los procesos de colisión con las partículas elementales de la materia, en eventos que emiten energía. El proceso inverso también sucede. Por ser difícil la ubicación de las partículas elementales en el tiempo y en el espacio a escala ultramicroscópico, la determinación exacta de la velocidad y la posición de las mismas, es sólo probabilística y es gobernada por el principio de la incertidumbre creada por Heisenberg en 1926 (Heisenberg, 1980: 11-78, Cassidy, 1992: 66-73). Física y filosóficamente, la mecánica cuántica, es en ciertas circunstancias: el estado de estar y no estar, ser y no ser al mismo tiempo. Lo que ha quedado perennizado en el clásico ejemplo del gato vivo - muerto, formulado por Schrödinger (Yam, 1997: 104-109).
 |
FOTOGRAFIA DE UN COMETA |
La teoría evolutiva del Universo expansivo está basada en dos hechos reales y un supuesto altamente probable, a saber:
- La comprobación persistente del corrimiento hacia el extremo rojo del espectro luminoso, de la luz emitida por la mayoría de las galaxias observadas. Las diferentes frecuencias de luz, son las que el ojo humano ve, como diferentes colores, correspondiendo las frecuencias más altas a los objetos más cercanos, que son identificados por el color azul en un extremo del espectro. Las frecuencias más bajas de los objetos más lejanos, corresponden al extremo rojo del espectro. El astrónomo Hubble en 1930, encontró que la mayoría de las galaxias que él estaba estudiando, tenían un corrimiento hacia el rojo, es decir se estaban alejando de nosotros, y este desplazamiento ni siquiera era aleatorio, era proporcional a la distancia que nos separa de ellas, es decir que cuando más lejos está una galaxia de nosotros, se aleja a mayor velocidad (Osterbrock et al.,1993:70-75). En otras palabras, el Universo continúa expandiéndose.
- Los primeros conglomerados estelares, formados a poco de haberse producido la gran explosión inicial, conllevaron a rápidos procesos de fusión nuclear para la formación de elementos. Lo reverso: la fisión nuclear producida poco después, en el interior de las estrellas emitió calor y radiaciones luminosas. La captación de éstas, en forma de luz (radiación de fondo), nos llega en forma de microondas, las que son captadas todo el ańo, en cualquier instante, ya sea de día o de noche. En 1965, dos físicos norteamericanos captaron esta radiación, cuando estaban experimentando con un detector de microondas extremadamente sensible. Y como la intensidad de las mismas no variaba, en cualquier dirección en que apuntara el detector, o cuales quiera que fuese la inclinación del eje terrestre, hizo suponer a Penzias y Wilson que provenía más allá del sistema solar, e incluso más allá de nuestra galaxia. Concluyeron que esta radiación era emitida por las estrellas más antiguas de las que se tenía conocimiento, había viajado a través de todo el universo observable, y nos continúa alcanzando hoy en día. Las características de esta radiación de ser siempre la misma en todas las direcciones en que se expanda (Hawking, 1992:67-75).
- El conocimiento de que ciertas estrellas que han agotado su combustible de hidrógeno colapsan su espacio, su volumen y su energía de modo al parecer infinito, estableciendo en su periferia un horizonte de sucesos, de los que incluso la misma luz no podría escapar, siempre y cuando hayan superado el límite de masa "normal", creado por Chandraseckar; ha originado el concepto de "agujeros negros" (Davies, 1986:39-42). La suma de las fuerzas gravitatorias de los agujeros negros, las estrellas, los planetas y otros, ejercería un efecto contrario a la natural expansión del Universo.
El análisis de las radiaciones de objetos luminosos muy distantes y cuasiestelares (quasars), ha permitido calcular que el Universo se originó probablemente hace 15,000 Ma, a partir de un suceso caracterizado por elevadísimas temperaturas, en los que existían estados extremadamente densos de energía y extremos de acción de las fuerzas gravitatorias y de la curvatura que éstos originan. Poco se sabe de los momentos previos a la gran expansión (big bang) porque la física y las matemáticas actuales no pueden mensurarla por el momento. Se dice que esto es así, porque las cantidades serían infinitas ( Hawking, 1992:76). Al respecto, varios científicos de nota, han expresado su acuerdo en el sentido de aceptar la existencia de una singularidad (región del espacio- tiempo, con curvaturas infinitas), previa a la gran explosión. En el otro extremo, Miró- Quesada afirma que el modelo standard de la gran explosión, al reducir el origen del universo a un punto pequeńísimo, con curvaturas extremas del espacio-tiempo, presenta la gran dificultad de manejar cantidades infinitas, que no tienen ningún sentido físico, y que por lo tanto deben ser descartadas (Miró - Quesada, 1991:441-45). Linde asimismo, opina que en tal singularidad sólo cabrían 10 partículas elementales, en comparación con las 1088 partículas elementales calculadas para el universo actual (Linde, 1995:17-20).
Producido el big bang y cuando la densidad del Universo en expansión cayó por debajo de la densidad de Planck: 1094 gr / cm3, y la edad del Universo era de 10-12 s, recién fue posible validar algunas leyes de la física actual (Linde,1995 :16-23). Pocos segundos después, cuando la temperatura descendió a 1015 grados centígrados, la materia y la energía adquirieron sus propiedades actuales, siendo ya posible entonces, realizar cálculos matemáticos con mayor precisión. Las partículas elementales navegaban en este momento en un mar de energía. Al expandirse el Universo unas mil veces más (10-5 s de edad ), toda la materia llenaba ya una región del tamańo del sistema solar. Tres minutos después, y de resultas de la colisión entre quarks y antiquarks, y de electrones y antielectrones, se logró finalmente obtener un discreto exceso de materia sobre antimateria (calculado en 1010) (Weinberg, 1994: 6-7). En esos momentos 3 quarks, se unieron para formar protones y / o neutrones. Finalmente, cuando el cosmos creció mil veces más (102 s de edad ), protones y neutrones se combinaron entre sí, formando núcleos atómicos elementales (Peebles et al., 1994: 13-15, Weinberg, 1994: 6-10). El predominio de los protones, determinó de los exceso de hidrógeno presente en el universo actual. Los modernos telescopios conectados a espectroscopios, siempre han detectado en los quasars, presencia de hidrógeno (H) y helio (He), con predominio del primero (Hogan, 1997: 10-16).
La explosión, al generar altísimas temperaturas, promovió, que la masa de energía adquiriera un color blanco incandescente, generando un inmenso resplandor, que aún no desaparece (radiación de fondo, adicional). Dicus afirma que la evolución ulterior del Universo fue determinada en los pocos momentos iniciales a las acaecencia del big bang. Así pues, la expansión continua del Universo constituye el paso inicial primordial de la evolución de la materia inorgánica (Dicus et a, 1983: 74-78).
Como ya se indicó esta evolución expansiva, tendría en la atracción gravitatoria de los planetas, agujeros negros, y otros conglomerados de materia aun desconocidos, su contraparte respectiva, las que de conjunto y en un momento determinado, podrían frenar o detener la expansión. Algunos creen que las galaxias sólo se están expandiendo para evitar colapsarse, posibilidad conocida como la gran implosión (big crunch) (Hawking, 1992: 26-27). Dicus y otros autores, que de producirse la gran implosión, ésta promovería ulteriormente expansiones cíclicas, cada vez más amplias.
La vida tal como la conocemos en el planeta Tierra sólo sería posible, durante las etapas expansivas del ciclo (Dicus et al., 1983: 84). A este respecto Linde opina que el modelo standard del Universo, con singularidades de por medio, no puede explicar las curvaturas infinitas, y en tal razón exige modelos más coherentes. El mismo sugiere la existencia de universos inflacionarios autorregenerantes infinitos, es decir de masas compactas de energía y materia llevadas a ese estado por implosiones previas, las mismas que transitando violentamente por túneles del espacio tiempo ("agujeros de gusano"), emergerían de novo en forma infinitos y autorrenovados big bangs. El universo actual sería sólo una de estas probabilísticas emergencias. Claro que este modelo necesita eso sí, de la existencia de muchos Universos (Linde, 1995:17-23).
La evolución del Universo estaría pues, en última instancia, determinado por el balance de las fuerzas expansivas y de las que frenan. El término medio habría ayudado a construir el universo actual. Una mayor compresión del sentido de esta evolución podría ser lograda, si se conociesen detalles previos al big bang (Haliwell, 1991:78-65).
La organización de los átomos de hidrógeno y helio, permitió ulteriormente la conformación de otros elementos por mecanismos de fusión nuclear. Así quedó constituida la Vía Láctea, la que se cree se originó por el colapso de nubes de gases, condicionadas por explosiones estelares previas, seguidas de la captura de fragmentos galácticos (Van den Bergh and Hessen, 1993:52-60).
 |
FOTOGRAFIA DE LA TIERRA |
Bajo este mismo modelo se formó la Tierra, a partir de polvo cósmico interestelar, el mismo que al inicio se agrupó formando partículas, las que devinieron en masas cada vez más grandes (crecimiento por acreción), hasta adquirir el tamańo de planetas pequeńos (planetesimales). El impacto frecuente de meteoritos, en esta masa inicial, logró la conformación de un núcleo central. Nuevas y adicionales acreciones, formaron en los siguientes 100Ma, el tamańo y forma final de la Tierra. Rocas halladas en los continentes (las únicas áreas de la corteza terrestre no sometidas al reciclaje rocoso), y datadas con Xe129, U238- Pb206, y U235-Pb207, han permitido fijar la edad de la Tierra en 4500 Ma (Allégre y Schneider, 1994 : 36-45).
Se desconoce la composición de la atmósfera en los primeros 200 Ma del universo en formación. Pero dataciones con Xe129 de gases atrapados en minerales dragados del suelo de los fondos marinos han permitido establecer con bastante certeza que la atmósfera primitiva consistía fundamentalmente de anhídrido carbónico (CO2), siendo el nitrógeno, el segundo componente más importante. (Mattioli and Wood, 1986:626-628). Existían también cantidades mínimas de metano (CH4), amoníaco (NH3), sulfuro de hidrógeno (H2S), ácido clorhídrico (HCl) y una cuasi ausencia total de oxígeno (O), tal como lo predijeran Oparin y Haldane a fines de 1920 (Oparín, 1979:15-26).
En términos comparativos, las atmósferas de planetas gigantes como Saturno y Neptuno, también son reductoras. El espectrofotómetro de onda corta ha puesto de manifiesto que la atmósfera de estos planetas está constituida fundamentalmente por hidrógeno, helio, y metano (Feuchtgruber et al., 1997. 159-164).
![]()
![]()
4.-Evolución de los elementos.
 |
Como ya se mencionó hace 15 000 Ma se produjo el fenómeno conocido como el big bang (la gran explosión). Cuando el Universo tenía 10-12s de edad, se pasó rápidamente del calentamiento inicial de la energía y la materia a un proceso de expansión y enfriamiento. Al expandirse el Universo hasta un 0,1% de su tamańo actual, y cuando la temperatura había descendido hasta 1015 grados centígrados, los quarks se condensaron creando protones y neutrones. Estos últimos al acoplarse entre sí formaron deuterones. Como los protones eran más numerosos que los neutrones, la mayoría de los primeros quedaron aislados, y se convirtieron en núcleos de hidrógeno. Los deuterones formados, se combinaron entre sí produciendo núcleos de helio, excepto una pequeńa proporcíón que hoy se pretende ubicar (Weinberg, 1994: 6-11). El hidrógeno (número atómico:1, peso atómico: 1,00), es el principal componente del polvo interestelar. De conjunto con otros gases y mediante procesos de condensación, ha contribuido a la formación de galaxias, estrellas y planetas (Hogan, 1997:10-14, Weinberg, 1994:7), El Universo considerado en su totalidad, está constituido esencialmente por hidrógeno (92,8%) y helio (7,1%) hallándose presentes como impurezas minoritarias: nitrógeno, oxígeno, neón y otros. La abundancia de un elemento disminuye en general al aumentar su número atómico, y los átomos de números atómico pares, son más abundantes (Dickerson, 1979:30-35).
Teniendo como base al hidrógeno (H) se formó el resto de los elementos, por procesos de fusión que liberaban energía. Hoy se sabe que en la zona central de las estrellas recién formadas (donde se han estudiado mejor estos procesos) existen temperaturas elevadísimas (del orden de 15 x 106 k). Allí los átomos de H (portadores de un protón), pierden sus electrones; los protones desnudos recorren el interior, experimentan violentas colisiones entre sí, siguen el curso de una serie de reacciones nucleares adicionales, se fusionan y producen núcleos de helio (He) , constituidos por dos protones y dos neutrones- densidad: 0,13, peso atómico: 4)], emitiendo energía, 2 positrones y 2 neutrinos (Kirshner, 1994:21-27).
Cuando la mayor parte del hidrógeno ha sido consumido, la conformación de la estrella se altera: sus capas externas se enfrían algo, se hinchan, adquieren gran luminosidad, y se convierten en estrellas gigantes de color rojo. Su núcleo por otro lado se contrae ganando en densidad y calor. En este estadío de madurez, los núcleos de estas estrellas promueven otro cambio evolutivo: contactos breves entre núcleos de helio producen fusiones. La colisión inicial de 2 núcleos de helio produce berilio (Be) (constituido por 4 protones y cuatro neutrones) que aunque muy inestables sirve para que al chocar, ocasionalmente con otro núcleo de helio genere un núcleo de carbono (C) (Hawking, 1992:116-119). Una ulterior fusión del helio con el carbono, originará oxígeno (O), litio (Li), berilio (Be) y boro (Bo), cuyos núcleos son más ligeros que el del C, aunque son también un millón de veces menos abundantes que éste. En realidad el carbono y el oxígeno son los elementos más numerosos fabricados en las estrellas (Arnett and Bazan, 1997:1359-1362, Weinberg, 1994:23). A este nivel, la escasez de los otros elementos cuyo núcleo tiene un par de protones. Esto explica la escasez de los otros elementos se debe a que en el Universo predominan los elementos cuyo núcleo tiene un par de protones. Esto explica la escasez del berilio porque la fusión de tres núcleos adicionales de helio, para la formación del carbono, está más favorecida que la reacción que se detiene después de la fusión de sólo dos núcleos de helio. Las estrellas medianas y pequeńas (estrellas solares) activas, sólo producen en general elementos ligeros, que promueven liberación de energía. El agotamiento del combustible nuclear de las mismas, originará estrellas enanas blancas (Kirshner, 1994:24).
No sucede lo mismo con estrellas de mayor masa que las solares. Aquí las reacciones termonucleares son transiciones efectivas de una vía secuencial predeterminada; que va "creando" nuevos elementos los que hacen un alto, luego que se ha formado el hierro. Cuando la estrella ha fabricado una zona central de hierro, cesa la generación de energía proveída por procesos de fusión nuclear. El núcleo del hierro, fuertemente ligado, tiene una energía total muy baja, por unidad de masa. Por esta razón algunos evolucionistas de la materia opinan cuando se ha llegado a este nivel, que "se llegó al límite del hierro" por que en este punto, cesa la generación de energía estelar, porque ha sido enteramente liberada (Dickerson, 1975:31-35).
En la fusión de los núcleos ligeros se desprende energía. Para fabricar elementos o núcleos de mayor masa que el hierro se necesita aporte y ganancia de energía. Por esa razón los elementos más pesados se forman en estrellas de masa enorme, mucho mayores que el sol, en las que se desarrollan procesos termonucleares de mayor amplitud. Aquí los núcleos de C, se fusionan creando neón (Ne) y magnesio (Mg). La fusión de los núcleos de oxígeno origina silicio (Si) y azufre (S). La fusión de los núcleos de silicio origina hierro (Arnett and Bazán, 1997:136). Cuando estas estrellas han formado su núcleo central de hierro, al no poder generar más energía: colapsan, su materia va alcanzando la densidad de los núcleos atómicos, y al no poderse comprimir más, pueden devenir en agujeros negros o en estrellas emisoras de neutrones. Si cae materia sobre este núcleo central duro, la onda rebota, retumba, se moviliza a velocidad supersónica a través de la estrella y al llegar a la superficie de la misma, despide un intensísimo brillo y explota. Estas explosiones denominadas de supernova II (Hawking, 1992: 116-119), generan más elementos, al eyectar helio a las capas más exteriores no consumidas aún, las que por su parte contienen carbono, azufre, silicio, oxígeno, y otros elementos que fueron acumulándose en sucesivas fusiones previas. En su avance a través de la estrella la onda de choque va provocando la síntesis de nuevos elementos, por el intenso calor que desencadenan las reacciones nucleares. Algunos de los productos de esta combustión son radioactivos. Se sintetizan otros elementos estables y más pesados que el hierro. Los neutrones bombardean los núcleos de hierro y los transforman en núcleos de oro. El oro se transmuta en plomo. El bombardeo nuclear del plomo genera el resto de los elementos hasta llegar al uranio (Kirshner, 1994:23-25). La fragmentación de parte de las capas externas de esta estrella explica la dispersión de los elementos en el espacio cósmico.
La Tierra consta de un núcleo de níquel y hierro y un manto que contiene olivina (FeMgSiO4). El carbono sólo constituye un 0,034% de la misma. Posteriormente la Tierra se estratificaría en un núcleo, un manto y una corteza, como consecuencia de la formación del planeta por acreción (Oparín,1979 :20-46).
Al principio la Tierra tenía temperaturas muy elevadas como para que el agua permaneciera en estado líquido. Cuando la temperatura descendió por debajo del punto de ebullición, el agua empezó a liberarse del interior ayudaba por procesos que liberaron gases, luego el agua se encauzó formando los oceanos primitivos. La emanación de gases formó una atmósfera secundaria compuesta por vapor de agua, NH3, CO2, CO, CH4, N, H2S y HCL (Allégre and Schneider, 1994:37-43).
Luego de la formación de la Tierra, la combinación de una gravedad baja y una elevada temperatura indujo a una pérdida de la mayoría de los elementos volátiles, los que se disiparon en el espacio interplanetario, poco después que el planeta se condensara alrededor de un centro. La abundancia relativa del oxígeno aumentó, porque quedó prisionero de minerales no volátiles como los silicatos, y porque oxidó a casi todos los minerales reducidos de los fondos marinos. Se perdió una gran cantidad de nitrógeno porque los nitruros son menos estables y más fácilmente transformables en gases volátiles (Dickerson, 1979: 34-36).
Varios de los elementos abundantes en el cuerpo humano son componentes de la atmósfera actual de la atmósfera primitiva ancestral y de los gases interestelar primigenios. Los 4 elementos más abundantes de los organismos vivos son el O, el H, el N, y el C, los que constituyen más del 99% de la masa de la mayor parte de las células. Como se ve, la evolución de los elementos ha favorecido a los más ligeros, para la conformación de la vida en el planeta Tierra. Ignoramos si en planetas con mayor masa y mayor gravedad de los fundamentos tengan que ser los mismos. Otras sustancias como el silicio también tienen capacidad para formar enlaces covalentes consigo mismo, y con otras sustancias, pero la oxidación rápida les impide formar estructuras (Lehninger 1993, 56-58).
![]()
![]()
5.-Evolución Química: El Carbono
El carbono es esencial para muchos procesos físicos y químicos que ocurren en la Tierra y en el Universo. Es uno de los elementos más abundantes en el espacio interestelar y en las estrellas. En las estrellas medianas (solares) se forma por la fusión del berilio y el helio. En el espacio interestelar gobierna procesos físicos y químicos, y probablemente forma la estructura del polvo interestelar, recursos importantes de la formación de los sistemas solares. El carbono es también el constituyente atmosférico predominante en planetas cercanos, como Venus y Marte (Snow and Witt, 1995:1455-1460).
El carbono puede compartir pares de electrones consigo mismo, formando enlaces C-C (carbono-carbono), de gran estabilidad. También puede formar enlaces simples y dobles con el oxígeno y el nitrógeno. Evidencia de ello es la conformación de monómeros simples, como las moléculas de ácido cianhídrico (HCN), cianógeno (C2N2) y compuestos con grupos CHO, los que constituyen pasos previos importantes hacia la síntesis de aminoácidos. Las rutas hipotéticas evolutivas de estas moléculas carbonadas simples en medios prebióticos ancestrales, encontrarían un adecuado respaldo en los experimentos de Miller. Este demostró que mezclas de hidrógeno (HCN), originando aminonitrilo (C2N2H4). Éste, al reaccionar consigo mismo y luego de eliminar dos moléculas de agua, conformaba finalmente glicina (Miller, 1953:528). Claro que en ausencia de oxígeno libre que pudiese degradar a los aminonitrilos, la hidrólisis pudo durar miles de ańos. En estas condiciones el formaldehído (CH3O) puede convertirse primero en ácido glicólico (C3H4O2) y luego en glicina. El acetaldehďdo (C2H3O) se convierte primero en ácido láctico y luego en alanina (C3H7NO2). Como se ve el carbono dominó la química prebiótica (Dickerson, 1979:91-173).
Las moléculas que tienen esqueletos carbonados unidos covalentemente se denominan compuestos orgánicos. El número de ellas y sus variedades son casi ilimitados, y están destinadas a la conformación de estructuras precursoras de la vida. La evolución ulterior del carbono tuvo como base tres tipos de reacciones iniciales de carácter fundamental:
- Alargamiento de las cadenas de átomos de carbono y el desarrollo del proceso inverso (ruptura de enlaces formados)
- Polimerización o combinación de dos o más moléculas orgánicas usando como puentes al nitrógeno, o al oxígeno, y el desarrollo del proceso inverso (hidrólisis) y
- Reacciones de óxido -reducción (Oparin,1979:91-173).
La mayor parte de las biomoléculas, son derivadas de los hidrocarburos (Dickerson, 1979:36-38). La evolución de los hidrocarburos, demandó a los átomos de hidrógeno su sustitución ulterior, por una amplia gama de grupos funcionales para originar familias de compuestos orgánicos con propiedades específicas. Los átomos de carbono unidos covalentemente pueden formar cadenas lineales ramificadas, estructuras cíclicas y en forma de celda, mayormente monocíclicas. En los seres vivos se conocen anillos de 5 aristas unidas a otras de 6, para formar compuestos aromáticos estables como los ácidos nucleicos: adenina y guanina (Curl y Smalley, 1991:14-20 ; Oparin, 1979 :20-71).
Para comprender la evolución ulterior del carbono, es necesario seguirlos fuera de los organismos vivos. Y aquí importa revisar conocimientos con relación a la conformación de ciertos compuestos carbonados policíclicos, grandes vacíos, tridimensionales y de forma de pelotas o esferas cerradas: los fullerenos (C60, C540, C960,etc). Estos, han sido obtenidos artificialmente evaporando barras de gráfito (primera forma del carbono, con uniones de tipo lineal), con láser, en atmósferas de helio (Curl y Smalley, 1991:14-24). Otros fullerenos, como el dodecabenzocoroneno, compuesto de 19 anilos bencenoides hexagonales, probablemente existe en las nebulosas (Aihara,1992: 62-64).
Sólo 30 elementos, son esenciales para los organismos vivos. La mayoría de los elementos constituyentes de la materia viva tienen números atómicos relativamente bajos. Cinco de ellos tienen números superiores al del selenio (Se). Los 4 elementos más abundantes en los seres vivos en términos porcentuales son el hidrógeno, el oxígeno, el carbono y el nitrógeno, que de conjunto representan más del 99% de la masa total, de la mayor parte de la célula. Estos elementos además de ser los más ligeros, tienen capacidad para formar enlaces estables (base para la formación de estructuras biológicas), supone una suerte de selección natural específica, por lo menos para el planeta en que habitamos. En la corteza terrestre, en términos porcentuales casi no existe carbono libre, en el agua de mar existe en una proporción de 0,0014%; mientras que en el ser humano alcanza la proporción de 9,5%. Como se ve, la química de los organismos vivos se organiza alrededor del elemento carbono (C), que representa más de la mitad del peso seco de las células. Los oligoelementos (Zn, Na, K, etc.) sólo representan una minúscula fracción del peso del cuerpo humano, sin que esto les reste importancia como elementos esenciales de la vida (Lehninger,1993: 18-32, 57-58).
La elección del carbono para formar estructuras biológicas, está basada en su gran estabilidad. Esto se debe a que mientras ciertos átomos, al ceder electrones, desprendiéndose de ellos, se convierten en iones negativos; al aceptar al incorporarlos, se convierte en un ion positivo. La unión de este tipo, define a los denominados enlaces iónicos. El carbono, que posee 4 electrones de valencia, en lugar de cederlos o aceptarlos, prefiere compartirlos mediante enlaces covalentes (Gribbin, 1986:169).
Para el caso de los enlaces simples C-C, o C-O, la fortaleza del enlace depende de la electronegatividad de los átomos enlazados. La electronegatividad en el caso C-C es nula, por lo tanto el enlace es muy estable. En el caso de elementos con diferente afinidad electrónica (electronegatividad) como C-O, el enlace se polariza, es decir que los electrones compartidos tendrán más probabilidad de encontrarse en la región del átomo más electronegativo, (en este caso el O), lo que hace al enlace, menos estable. El número de electrones compartidos y la distancia de los de enlace, con respecto a cada núcleo también afecta la fuerza del enlace. Los enlaces carbonados dobles son más fuertes que los simples, y los triples, lo son más que los dobles. Ningún otro elemento puede constituir moléculas con formas y tamańos tan diferentes y con tanta variedad de grupos funcionales como el carbono (Lehninger, 1993:58-59).
Con relación a enlaces carbonados dobles, importantes aportes, en relación con la estabilidad de los mismos, han sido proporcionados por estudios realizados en el benceno y los fullerenos. Del benceno se sabe que su extraordinaria estabilidad reside en el hecho de que todos sus átomos de carbono presentan órbitas llenas de electrones. En este caso, cada átomo de carbono comparte más de dos electrones de valencia con el átomo de carbono más cercano. Las dos uniones que integran los enlaces dobles del benceno, se conocen con los nombres de enlace sigma, (enlace sencillo y fuerte) y enlace pi (al parecer inestable y débil en apariencia). Los orbitales atómicos, en los que residen los electrones pi, al ser deslocalizados, se solapan con los de sus vecinos y se movilizan por zonas comunes formando nubes pi, en forma de rosquillas. Esta características de viajar por orbitales comunes contribuye a la estabilidad del enlace. Si el número de electrones pi, que conforman los orbitales externos, es: 2, 6, 10, 14 etc, los enlaces serán fuertes, lo que a su vez determinará acopio de energía en cantidades que pueden ser predichas (Aihara,1992:62-66). El hecho de que este aserto falle en los fullerenos, se debe a que las estructuras cíclicas incorporan una energía adicional, positiva (energía de resonancia topológica, no presente en las estructuras carbonadas lineales), y que al parecer es promovida por la propia estructura tridimensional del compuesto. Cálculos matemáticos topológicos, resumidos en la ecuación de un polinomio característico, creado por Hosoya y Aihara, han acercado bastante los valores de energía calculados, para romper cierto número de enlaces carbonados (energía de atomización), que nos permite predecir qué compuesto tendrá un enlace fuerte. En general todos los compuestos aromáticos con energía positiva, presente aún en algunos compuestos carbonados del tipo de los fullerenos, sólo indica nuestra precariedad de conocimientos con relación a los fundamentos últimos condicionadores de los fuertes enlaces del carbono (Aihara and Hosoya, 1988:2657-2659).
Así pues la evolución estructural del carbono en los seres vivos se inicia con la creación de cadenas lineales, covalentes (como las largas uniones de los ácidos grasos), las mismas que llegado a un punto, devienen en anillamientos del tipo del colesterol y los pigmentos de las plantas (Schopf, 1979:56-57). Al final y según la propuesta de Ingber, existirían reglas universales que parecerían guiar el diseńo de las estructuras orgánicas, desde elementales compuestos carbonados hasta células complejas y complicados tejidos y organismos (Ingber, 1998:30). Las proteínas y los polinucleótidos tienen también tendencias a anillarse en forma reversible y espontánea, efecto que parece depender de las uniones hidrogenadas y de demostrar que compuestos hidrocarbonados artificiales, tornan sus estructuras lineales en helicoidales, han asumido que estas conformaciones son promovidas casi exclusivamente por enlaces carbonados (Nelson, 1997:1793-1796). Los radiolarios, protozoos de esqueleto silíceo, desarrollan estructuras fullerénicas, al igual que ciertos virus, organelas y ciertas enzimas celulares (Curl and Smalley, 1991:23, Ingber, 1998:31-39).
![]()
![]()
6.-Evolución Química: Monomeros
Esta es un área de la historia evolutiva con grandes vacíos y escasas evidencias, pero con significativas construcciones teóricas que sirven de guías a diversos experimentos. Leamos reflexivamente la siguiente panorámica:
Formación no biológica de monómeros:
La mayoría de las moléculas sencillas, o monómeros: bases púricas o pirimídicas, ribosa o desoxiribosa y aminoácidos, relacionadas con procesos vitales y necesarios para la construcción de los sistemas genéticos de organismos vivos ancestrales, son posibles de ser sintetizadas en condiciones simuladas de la Tierra primitiva (Orgel, 1979:10-95).
La formación de monómeros es posible en atmósferas artificiales que consista de gases como hidrógeno (H); y de sus formas totalmente reducidas: amoníaco (NH3), metano (CH4), y agua (H2O). El primer paso observable en estas condiciones es la constante reducción en la concentración de NH3 para dar lugar a estructuras monoméricas simples como el ácido cianhídrico (HCN), cianógeno (C2N2), compuestos con el grupo CHO y bases púricas y pirimídicas. Todos estos gases circulando por 7 días, en un sistema que emite chispas eléctricas (sustituido incluso por rayos ultravioleta), produce también con mucha facilidad muchos aminoácidos típicos: glicina, alanina, isovalina, prolina, sarcosina, beta - alanina, ácidos alfa y beta aminobutírico, norvalina, isovalina, prolina, ácido aspártico, norleucina, ácido pipecólico y otros (Miller, 1953:528-529). La formación de monómeros se logra también en atmósferas prebióticas más simples,como las conformadas por CO2, nitrógeno, agua y trazas de hidrógeno (Mattioli and Wood, 1986:626). Los nucleótidos son formados con facilidad en atmósferas reductoras (ricas en hidrógeno). En atmósferas oxidantes esta posibilidad es mínima, porque el oxígeno captura átomos de hidrógeno de otros compuestos, impidiendo reacciones que permiten transformar moléculas orgánicas sencillas, en otras más complejas (Oparin, 1979:122-173).
Algunas de estas unidades monoméricas son susceptibles de ensamblarse, para constituir 2 grandes clases de polímeros: ácidos nucleicos y proteínas (Orgel, 1994:76-80).
Monómeros importantes para la formación de los ácidos nucleicos son dos tipos de azúcares: ribosa (para el RNA) y desoxiribosa (para el DNA). Cuesta sintetizar ribosa en cantidad y pureza adecuada en ausencia de enzimas, porque las reacciones conducentes a ello, crean una mezcla de azúcares, en las que la ribosa es siempre un componente minoritario. Asimismo los demás azúcares, combinados con bases nucleicas circundantes, tienden a formar productos que inhiben la síntesis y la replicación del RNA (Orgel,1994:76-77).
Hoy en día se acepta que estos azucares podrían haberse formado, en condiciones prebióticas a través de una reacción de varios pasos, que implica la condensación de 5 moleculas de formaldehído para originar una molécula de ribosa (reacción de la formosa). Aunque a decir verdad esta reacción, proporciona muchos azúcares, sin ninguna selectividad (Dickerson, 1979: 37-46). De otro lado Larralde y Miller, han demostrado que la ribosa y otros azúcares en condiciones ancestrales, son inestables en presencia de ácidos o álcalis fuertes, por lo que han sugerido examinar estructuras de soporte prebióticas, no basadas en azúcares (como los péptidos de ácidos nucleicos), que podrían haberlas precedido. Admiten sin embargo que la elección de la ribosa podría haberse dado en condiciones especiales, basada en procesos de selección molecular (Larralde et al., 1995:81-58). Erschenmoser ha tratado de obviar esta limitación produciendo casi exclusivamente ribosa, a partir de la polimerización de moléculas de formaldehído, con pasos intermedios, que requirieron previamente la transposición de grupos fosfato, para que el producto principal fuese un intermediario fosforilado de la ribosa (Horgan, 1991:119).
Otro básico, quizás, el más importante componente del código genético es la adenina. Aunque Juan de Orá obtuvo adenina y otros aminoácidos a partir de soluciones acuosas de HCN y NH3, sin ayuda de aldehídos (Oró and Kimball, 1961:217-227), es también probable que la formación inicial de la adenina tuviese que ver con la combinación de 4 moléculas de HCN, formando inicialmente un tetrámero de HCN, el diaminomaloniltrilo: (HCN)4 (importante intermediario en muchas reacciones que conducen a la síntesis de bases). El tetrámero en presencia de luz, habría sufrido un proceso de autoreordenación que lo habría capacitado para captar otra molécula de HCN, constituyendo finalmente un pentámero de HCN: la adenina. Esta síntesis pudo efectuarse en condiciones similares a los de la Tierra primitiva, porque esta base purínica es fácil de sintetizar. Existió selección molecular aquí, porque de 3 isómeros posibles sólo la adenina fue incorporada a los seres vivos (Dickerson, 1979:36-42). Por reacciones de hidrólisis que comprometen al diaminomalonilnitrilo, y en las que interviene el cianógeno (C2N2), se pudo formar la otra purina. La guanina. Las restantes bases de ácidos nucleicos: timina (T); uridina (U) y citosina (C), pueden obtenerse a partir de reacciones entre cianuro de hidrógeno y otros dos componentes : cianógeno (C2N2) y cianacetileno (C3HN), capaces de ser formados en atmósferas prebióticas reductoras (Orgel, 1994: 78-60).
La adenina y la guanina han sido halladas en el polvo interestelar, en áreas cercanas a las estrellas en formación y en meteoritos carbonáceos (como el meteorito de Murchinson), de conjunto con otros constituyentes elementales como formaldehído, agua, HCN, cianacetileno (C3HN), NH3 y otros (Cronin and Pizarello, 1997: 951-955, Zahnle and Greenspoon, 1990:157-160). Se ha hallado glicina en nubes, en vías de formar estrellas nuevas, cerca del centro de nuestra galaxia. De otro lado se han identificado también, hidrocarburos complejos en el interior del polvo interestelar microscópico (Clemett et al., 1993: 721-724).
La unión universal en el RNA y DNA, de la ribosa o de la desoxiribosa, con la adenina para formar un nucleósido de adenina, siempre se realiza a nivel de la configuración beta -1’ (configuración levógira). Esto implica un poderoso proceso de selección porque la ribosa tiene 4 carbonos (1’, 2’, 3’ y 5’) con grupos hidroxilo (OH), cualquiera de los cuales puede servir para el anclaje de la denina. Además tres de estos grupos, se hallan unidos a "carbonos asimétricos": 1’, 2’ y 3’ (carbonos posibles de promover configuraciones diferentes, o de estereoisómería ) (Dickerson,1979:40).
En las moléculas orgánicas se pueden formar estereoisómeros (moléculas con 2 configuraciones especulares D: dextrógiro y L: levógiro), cuando tienen 4 átomos diferentes o grupos diferentes unidos a estos átomos, como sucede frecuentemente con los átomos de carbono (C). En condiciones experimentales siempre se producen cantidades iguales de ambas formas ópticamente activas. Curiosamente, todos los organismos vivos tienen mayoritariamente configuraciones levógiras en sus estructuras (con la discreta excepción de las paredes bacterianas y los mecanismos de defensa bioquímica). Es decir que existe un favorecimiento o selección natural para uno sólo, de los isómeros ópticos. Al respecto Hegstrom y Kondepudi opinan que la homoquiralidad de las moléculas, con toda seguridad surgió antes de la emergencia de la vida misma, y fue una condición para la emergencia de la misma (Hegstrom and Kondepudi, 1990:58-69). Dickerson ańade que la selección de la unión beta -1’ entre monómeros, estuvo basada en la mayor eficacia funcional de las enzimas, cuando se unen a la superficie de un solo isómero (Dickerson, 1979:43-45).
Según Orgel, la obtención experimental de nucleótidos, ofrece al mismo tiempo muchas conformaciones incorrectas, lo que abona a favor de la existencia de procesos de selección de naturaleza molecular (Orgel,1994:78). La fuerza de estos procesos selectivos se hace evidente también, cuando uno cae en la cuenta de que sólo las purinas, y no las pirimidinas, pueden unirse a la ribosa (Dickerson, 1979:37-42).
Aunque en pequeńas cantidades, el fósforo inorgánico se hallaba presente en la Tierra primitiva y fue sin duda utilizado como fuente energética. Existe también la posibilidad de que moléculas de ácido alkilfosfónico soluble en agua, encontrado recientemente en meteoritos y cometas, hubiese aportado el suministro orgánico faltante (De Graaf et al., 1995:474-477). Orgel y colaboradores han producido nucleósido (ribosa unida a bases púricas), pero sin el grupo fosfato (Orgel 1994:51).
Cuando la adenina (A) se une a una molécula de ribosa, se forma un nucleósido, y si a éste, de modo sencillo se le agrega una cola de trifosfato se convierte en trifosfato de adenosina (ATP). El que los organismos vivos hayan escogido para intercambiar o almacenar energía al ATP, en lugar de GTP, CTP; o UTP, y emplear adenosina en lugar de guanosina constituye sin duda un proceso de selección (inducidos por estar presentes en mayor concentración y ser más fáciles de sintetizar). La unión prebiótica de fosfatos, ribosa y bases púricas y pirimídicas conformando nucleótidos fue la base para la construcción de los ácidos nucleicos. Ahora faltaba unirlos. El paso acoplador de monómeros (polimerización), necesita de 3 instancias 1)eliminación de moléculas de agua 2) medio de soporte para la polimerización: superficies de arcilla, por ejemplo, y 3) agentes acopladores, dadores de energía (Dickerson,1979:40).
![]()
![]()
7.-Formación de polímeros.
 |
Inicialmente se habría formado varios polímeros, uno solo de los cuales conseguiría autoreplicarse en forma correcta. Esta operación aunque es algo más complicada de lo que parece a simple vista, es perfectamente posible de llevarse a cabo en atmósferas primitivas (Orgel, 1994:77-84).
Para que los polímeros se unan en forma no biológica tiene que existir una unión previa, correcta entre nucleótidos, exigencia mayor que la requerida para las proteínas. La unión experimental más favorecida entre nucleótidos, aunque menos estable, desde el punto de vista químico es la 5’ -2’. Sin embargo, la unión universal actual entre la ribosa y la adenina tanto del RNA como del DNA, de los seres vivos, es la 5’, -3’ explicable sólo por la fuerte presión selectiva derivada de las grandes ventajas que esta unión da, para el almacenamiento de la información genética, dada su gran estabilidad. El largo brazo de las fuerzas selectivas, le ha exigido también al DNA, la eliminación del grupo hidroxilo -2’, para hacerlo más estable (Dickerson, 1979:40 48).
Según Orgel es difícil entender hoy en día, de qué modo las reacciones químicas de los tiempos prebióticos desembocaron en la conformación de ribonucleótidos necesario para establecer la autoreplicación del RNA (ácido ribonucleico) (Orgel, 1994: 82-83). Y se piensa que las respuestas a estas interrogantes deben yacer en las fuentes y en los modos energénicos.
Aunque la fuente de energía acopladora actual proviene del ATP, los primeros agentes acopladores inorgánicos, que promovieron la polimerización, quizás hayan sido las carbodiimidas, el cianógeno, la cianamida, el cianacetileno y el diaminomalonitrilo. Estos agentes con toda probabilidad promovieron inicialmente uniones incorrectas, produciendo moléculas emparentadas con nucleótidos que pudieron haber impedido incluso, la generación de más RNA. Por esa razón quizás haya sido necesaria la eficaz acción acopladora proporcionada por los sistemas genéticos sean viables. Ferris ha producido la síntesis prebiótica de oligómeros de RNA (de hasta 55 monómeros de longitud), en superficies de arcilla de montmorinollita; y de aminoácidos en arcillas de illita e hidroxiapatita (Ferris, 1996:59-61).
Según Cairns-Smith, es bastante factible que estos oligómeros hayan originado sistemas genéticos primitivos, los que en un inicio fueron elementales e inorgánicos. En su opinión ciertas irregularidades de la estructura de una arcilla (una distribución irregular de cationes por ejemplo), las hizo depositarias de información genética, y la replicación podría haberse realizado, si una disposición determinada de cationes de una capa de esta arcilla hubiese dirigido la síntesis de una nueva capa. La selección ulterior de una determinada capa, se hizo en función de su eficacia para el copiado. (Cairns-Smith, 1982:25-75). Como este sistema no ha sido engullido por formas acopladoras más avanzadas, las ideas en torno a sistemas genéticos ancestrales se movilizan hoy día, alrededor del más visible y cuantificable "Mundo del RNA".
El consenso actual sugiere que como los agentes acopladores primitivos fueron susceptibles de un agotamiento rápido, con toda seguridad fueron reemplazados, por la acción acopladora que a modo de protoenzimas, empezaron a desarrollar las propias cadenas poliméricas de Rna en formación. Algunos procesos de polimerización tienden a ser autocatalíticos: la presencia de un polímero determinado favorece la formación de más polímeros (Czostak, 1993:511-533). Si un sistema no es autocatalítico debe enfrentarser tarde o temprano a la merma del suministro de los agentes acopladores. Una enorme presión selectiva debió darse para crear un sistema económico y eficiente como el que se da en el altamente autocatalítico del DNA, para eliminar esta desventaja (Dickerson, 1979:41-42).
El DNA (ácido desoxiribonucleico) se construye a partir de 4 clases de unidades monoméricas: los desoxiribonucleótidos, y el RNA a partir de 4 tipos de ribonucleótidos. Sumados, estos polímeros lineales, dan 8 tipos de nucleótidos, a partir de los cuales se construyen todos los ácidos nucleicos, y los 20 diferentes tipos de aminoácidos, a partir de los cuales se forman las proteínas, que por otro lado son idénticas en todos los seres vivos (Lehninger, 1993:6). Aquí es necesario recalcar que existió selección natural, cuando de un número más vasto, se escogieron 20 alfa-aminoácidos, denominados primordiales o esenciales. Del mismo modo, hay selección cuando los organismos vivos deciden incorporar a la mayoría de sus estructuras sólo L-aminoácidos (Hegstrom and Kondepudi, 190.58-68, Engel and Macko, 1997:265-267). Con 20 aminoácidos, ya hay sillares suficientes para la conformación de proteínas. La energía para unir los aminoácidos, pudo provenir de protoenzimas como las carbodiimidas (N=C=N), poseedoras de enlaces de gran energía, o de cianógeno y cianamida (N=C-NH2). La carbodiimida se pone en contacto sucesivamente con dos monómeros o polímeros de los cuales, uno tiene un grupo OH y el otro un H terminal. El compuesto elimina agua, uniendo los dos monómeros o polímeros por sus extremos. La captación de agua por la carbodiimida libera energía y así continúa la reacción en forma combinada y cíclica (Dickerson, 1979:35-41). Recientemente, Rihe y Orgel han demostrado que la acilación oxidativa en soluciones, es un buen método para activar y condensar ácidos carboxílicos, incluyendo aminoácidos (Rihe and Orgel, 1997:52).
![]()
![]()
8.-Evolución Química: "El mundo del RNA"
 |
Como es difícil constituir polímeros sin la ayuda de enzimas, esta misma limitación apoya la posibilidad de que el RNA ancestral tuviese que ser autoreplicante (Orgel, 1994:77-82).Cech y Czostak han logrado acoplar secuencias cortas de nucleótidos, luego de modificar ribozimas naturales (Czostak, 1993:511-533). Joyce por métodos más refinados ha logrado que ribozimas naturales, rompan enlaces químicos de nucleótidos y péptidos (Joyce, 1989:83-87). Weinstein y colaboradores han demostrado que las ribozimas del grupo I intron de Tetrahymera termophyla, con la ayuda de iones divalentes como Mg"+, Mn2+ o Cd 2 + promuevan acciones autocatalíticas, y concomitante ligazón de exones en la guanosina (Weinstein et al., 1997:805). Spiegelman ha demostrado que si se deja replicar repetidamente RNA (ácido ribonucleico), en presencia de una ribonucleasa, ésta deviene en resistente a dicha enzima (Horgan, 1991: 116-125). Así, el descubrimiento de las propiedades autocatalíticas del RNA demostró que una sola estructura química puede funcionar como genoma y enzima, porque ciertos tipos de RNA (aunque con dificultad), actúan como sus propias enzima: se cortan en 2, y luego se acoplan (Cech, a987:1536-1537).
Estos hallazgos han permitido objetivar un "mundo del RNA", el cual habría seguido los siguientes pasos:
- Conformación del RNA, a partir de la ribosa y otros compuestos orgánicos
- El RNA evoluciona y se copia a sí mismo
- El RNA sintetiza proteínas que funcionan como catalizadores
- Las proteínas ayudan al replicado del RNA y a sintetizar proteínas en forma más eficiente. La doble cadena de RNA evoluciona y se convierte en DNA
- El DNA creado toma el comando y las funciones se dividen a) el RNA se dedica a fabricar proteínas y ayuda al DNA a hacer copias de si mismo y b) el DNA almacena la información genética, actúa de molde del RNA y se comporta como su propio autocatalizador (Gilbert, 1986:618, Tarasow et al., 1997: 54-57).
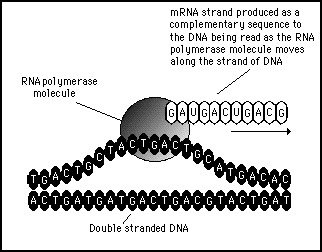 |
Por otro lado Erschenmoser ha demostrado con la creación de un piranosil-RNA ( p-Rna), que consta de una ribosa con un carbono adicional, que no es necesario el enrolamiento de la hélice bicatenaria para formar cadenas complementarias (Horgan, 1991 .120-125). Este hecho y la dificultad para obtener polimerizaciones a partir de componentes de RNA ha llevado a pensar en la existencia de sistemas genéticos (ancestrales) diferentes, con componentes más fáciles de sintetizar, como el ácido péptido nucleico (PNA). El ácido péptido nucleico semeja al RNA en su habilidad para formar complejos de doble hélice. El PNA, es sostenido por uniones de tipo amida (mas bien, del tipo fosfodiéster), pudiendo ocurrir transiciones de un sistema genético a otro, sin pérdida de la información (Böhler,1995.578-581). Artificios químicos le han permitido a Rebek autoreplicar moléculas orgánicas, como el éster triacídico de aminoadenosina -AATE-, una mezcla artificial parecida a nucleósido. El autor asumió que la capacidad autoreplicativa las tienen las moléculas que tienen forma y estructura química complementaria (Rebeck, 1994:48-55). Orgel y Joyce critican esta presunción por la exactitud de las copias, lo que revela que no hubo mutaciones. Según Joyce, el RNA, aprende a copiarse a si mismo en un proceso, sin ayuda de enzimas (Joyce, 1989:83-87). Ultimos experimentos de Joyce han demostrado que una población de moléculas naturales de RNA catalizadoras de sustratos templados de RNA, lograban al cabo de 52 horas una amplificación se habría llevado a cabo por procesos de amplificación y mutación (Wright and Joyce, 1997:614-617).trabajos recientes de Orgel han establecido que una vez creados los oligonúcleotidos, estos sólo se unen con sus pares de bases, denominados también- pares de base de Watson-Crick-: adenina con timina, guanina con citocina en el DNA; y adenina con uracilo en el RNA. Es decir que la formación de las cadenas complementarias sigue la ley de la atracción mutua, donde se establecen una vez más los criterios de la selección, y el rol de la quiralidad. Esta selección puede producirse por medios abiológicos aunque las enzimas tornan estos pasos más eficaces. Orgel ha enfatizado que la producción experimental de cadenas complementarias adicionales, sólo cuando se emplearon moldes dextrógiros. Cuando se emplearon cantidades iguales de foremas L y D, la producción se bloqueó (Orgel, 1994.77-85).
Así, la cooperación e interacción, entre proteínas (elementos de estructura y protección) y ácidos nucleicos, (autocatalizadores y moldeadores genéticos) se dio desde el inicio.
 |
|
VIRUS DE LA GRIPE |
VIRUS FAGO T4 |

Cuando las circunstancias exigieron economizar la energía, en proceso de extinción, las condiciones para evolucionar hacia unidades independientes, cuasi replicantes, ya estaban dadas. Así probablomente emergieron los virus, como complejos supramoleculares que pueden autoreplicarse, en células huésped adecuadas. Constan de una molécula de ácido nucleico: DNA o RNA, rodeada de una envoltura protectora (cápside). Fuera de la célula huésped carecen de vida y pueden cristalizar. Pero adoptan otros estadíos, cuando ellos o sus ácidos nucleicos penetran en las células y se convierten en parásitos intercelulares,al usar los ribosomas y las enzimas del huésped para producir otros viriones (Lehninger, 1993:50). La diversidad genómica de los virus contemporáneos, va desde la conformación estructural de una, o doble cadena de RNA o DNA, virus de DNA, virus de RNA, de RNA que replican a través de intermediarios de DNA, virus hechos de RNA y DNA, virus lineales o circulares. Maizels y Weiner sugieren una aparición evolutiva temprana de los virus, ya que su diversidad genómica refleja la variedad de las estrategias replicativas disponibles. Agregan los mismos autores que el análisis del genoma del virus QBRNA, al poner en evidencia factores replicantes y transcriptores, supone relaciones de larga data entre estas estructuras (Maizels and Weiner, 1994:6729-6734).
![]()
![]()
9.-Protobiontes.
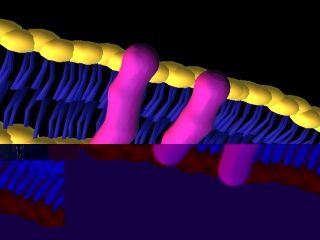 |
MEMBRANA CELULAR |
Era importante luego, dotar de una cubierta protectora al primitivo sistema replicador y aislarla del medio exterior. Estos mecanismos han sido estudiados al comprobar que mezclas secas de aminoácidos puros polimerizan espontáneamente; y a temperaturas no muy altas: 130°C, dan lugar a mezclas gelatinosas (constituidas de hasta 200 unidades aminoacídicas) denominadas "proteinoides termales". Según Fox y Harada, si estos proteinoides se hubiesen producido en medios prebióticos, se hubiesen arrastrado posteriormente hasta las escorias volcánicas, secándose por eváporación (polimerizándose, más aún), regresando finalmente a la superficie del mar, para volver a ser utilizados en algún momento posterior. Según Fox, estos proteinoides fueron los predecesores de la vida (Fox and Harada, 1960: 3745-3751).
|
OPARIN |

Trabajos independientes de Oparín, indican que los polímeros obtenidos de diversas formas, inmersos en soluciones acuosas tienden a separarse espontáneamente, paran formar coacervatos (gotitas con polímeros en su interior), suspendidos en un medio acuoso. La estabilización de los coacervatos depende de la adquisición de un metabolismo primitivo. Así, si se ańade fosforilasa a una solución de que contiene histonas y goma arábiga, la enzima queda concentrada en el interior de las gotitas, y si al agua del medio circundante se le ańade glucosa - 1-P, ésta difunde al interior de las gotitas polimerizándose por acción de la enzima, formando almidón. Si los coacervatos son muy grandes, se rompen y dan lugar a gotitas hijas. Los coacervatos serían ejemplos de protoorganismos (protobiontes), capaces de autoperpetuarse (Oparin, 1979:70-78).
Cuando los proteinoides de Fox, se calientan en soluciones acuosas a 130°C - 180°C, se agregan espontáneamente, formando microesferas de 1- 2 micras de diametro, que desarrollan delimitaciones externas parecidas a una doble capa lipídica. Es posible pues, que los primeros protobiontes fuesen coacervatos autocatalíticos, que dependían de agentes acopladores y monómeros activados, en lugar de enzimas. Con leves disensiones hoy en día, se acepta que las propias cadenas poliméricas de RNA en formación, fueron los primeros agentes acopladores, formándose así los primeros sistemas genéticos, no visibles actualmente, porque fueron eliminados por la selección natural (Gribbin, 1986:180-190).
Varios modelos teóricos tratan de explicar la forma en que ciertas superficies sólidas proporcionarían energía cíclica promoviendo el paso de protobiontes a arqueobacterias. De Duve y Miller sostienen que la energía fue proporcionada por tioésteres, y la nutrición por mecanismos heterótrofos (De Duve and Miller, 1991: 10014-10071). En el otro extremo y en energía fue proporcionada por pirita (sulfuro de hierro o cobre, presente en forma de cristales), existente en forma abundante en superficies sólidas. La pirita al ofrecer una superficie cargada positivamente, permitiría la unión de compuestos orgánicos. Wächtershäuser refuta la posición De Duve y Miller, porque la propuesta de ellos involucra demasiados pasos. Alternativamente sostiene que la nutrición ancestral se obtuvo por mecanismos quimioautotróficas (síntesis de parte o la totalidad de las unidades monoméricas y macromoléculas a partir de CO2 y NH3 utilizando los electrones de la luz solar). Plantea asimismo que en algún momento en cristal con polímeros en su interior rezumó otro cristal promoviéndose de este modo el surgimiento de la vida (Wächtershäuser, 1994: 4283-4287).
|
ARCHEOBACTERIA |
Cierto grado de relación entre lo precedente y células ancestrales, pretenden darle Farina y colaboradores, a los resultados de sus estudios sobre procariotas anaeróbicas del tipo de las arqueobacterias y el Aquaspirillum magnetotacticum, hallados en sedimentos acuosos. Ellos se dieron con la sorpresa de que éstas incorporan cristales de magnetita sulfuro de hierro, pirita y otros compuestos inorgánicos a su citoplasma celular. Estos compuestos inorgánicos si bien les ayudan en su orientación en el campo geomagnético, podrían también jugar un rol como fuentes de energía autotrófica (Farina et al., 1990:70-78). De otro lado Ohmoto y colaboradores, han sugerido que la pirita, hallada en los cinturones de Barbeton, Sudáfrica (3,4 Ma) fue formada por la reducción bacteriana del sulfato del agua de mar (Ohmoto et al., 1993;555).
![]()
![]()
10.-Evolución Celular.
 |
CELULAS BACTERIANAS |
De aldehídos y aminoácidos formados abiológicamente, en atmósferas ancestrales simuladas (Miller, 1953: 528) o engentradas en forma natural (meteorito carbonáceo de Murchinson), polvo interestelar), Hasta la emergencia de las primeras células; existen vacíos reproducibles e irreproducibles. Lo irreproducible, desaparecido por la competencia y la selección natural, es escrutado hoy por la analogía y los diseńos experimentales tendientes a certificar múltiples hipótesis (Horgan, 1991:119-122). Los reproducibles están basados en un magro registro de fósiles bacterianos, la historia geológica del planeta, el análisis de la filogenia de nucleótidos de arqueobacterias, procariotas y eubacterias y el escrutinio del material orgánico procedente del espacio interestelar (Schopf, 1979:51).
 |
 |
ALGAS |
BACTERIAS |
Según Woese, la última célula ancestral común, a todos los seres vivos fue un progenote: una estructura cuasi celular, poseedora de un elemental sistema de transferencia de información genética. Este progenote cuya estructura básica se fue delineando a través de un proceso de depuración de varias formas similares ancestrales ,se constituía de ribosomas sencillos. Su constitución implicaba sin embargo, un acoplamiento fenotipo-genotipo, ya cercano al de las bacterias actuales, lo que indicaba que continuaba evolucionando activamente en el sentido darwiniano (Woese, 1977:5088-5090, Doolittle and Brown, 1994; 6721-6728). Muchas evidencias sugieren que los progenotes, al evolucionar devinieron en procariotas ancestrales (Schopf, 1979:51-54) y desarrollaron en su interior estructuras del tipo de los peroxisomas (ejecutores de variadas funciones metabólicas), mitocondrias (generadoras de energía), y plástidos (lugares de realización de la fotosíntesis) (De Duve, 1996:38-40).
Las arqueobacterias, de las que se cree que son las células más antiguas (~ 4100 Ma), fueron estudiadas entre 1996 a 1976, mediante el secuenciamiento de sus núcleotidos rRNAs (ribosomas RNA). Las arqueobacterias viven en medios extremos /medios sulfurosos, salinos, ácidos, cercanía a fuentes hidrotermales, áreas de minerales calientes. etc).
Las más antiguas han podido sobrevivir en medios extremos de temperatura: 150°C (bacterias hipertermófilas) (Madigan and Marrs, 1997: 66-71). Los estudios filogéneticos de sus nucleótidos han permitido corroborar que las arqueobacterias tienen secuencias mínimas compartidas con eucariotas y eubacterias (Woese, 1977:5088-5090). Sin embargo, y como ha quedado comprobado con el secuenciamiento total de nucleótidos de Metanococcus janaschii ( arqueobacteria que emite metano como producto colateral), lo usual más bien, es que esta secuencia difiera en la mayor parte de las veces, con las que la mayoría de las otra bacterias tienen (Pace, 1997:734-740). La cercanía de la emergencia de estas bacterias, con un genoma consolidado, a la edad de la constitución inicial de la Tierra, ha instalado a algunos, medio en serio medio en broma, a plantearse la posibilidad de que éstas, pudiesen tener un origen extraterrestre (Orgel,1994:77-84).
[Componente Mapa de imágenes de FrontPage]
Las otras procariotas están representadas por cianobacterias (antes llamadas algas verde-azules) y otras, del tipo de Escherichia coli. Tienen un tamańo que oscila entre 1-10 micrómetros llegando raramente a los 60 micrómetros, metabolismo por fotosíntesis aeróbica o anaeróbica, son inmóviles o poseen flagelos, DNA circular en el citoplasma, reproducción binaria simple, paredes celulares compuestas de azúcares y péptidos característicos, presencia de orgánulos sin membranas, y conformación unicelular. Estas, a diferencia de las arqueobacterias habitan en el suelo, en las aguas superficiales, en los tejidos de ciertos organismos vivos, o en descomposición (Schopf,1979:53). El hallazgo de inclusiones carbonáceas (determinado como carbono ligero isotópico), en el interior de gránulos de apatita (fosfato cálcico básico), como evidencia de la existencia de vías biogénicas, ha permitido alargar la emergencia de procariotas de este tipo a alrededor de 3800 Ma (Mojzsis et al., 1996:55-69).
| [Componente Mapa de imágenes de FrontPage] | [Componente Mapa de imágenes de FrontPage] |
La presencia en mitocondrias y plástidos de sistemas genéticos vestigiales y funcionales, prueba que éstas, estaban contenidas en varios modelos de células primitivas de vida libre, las que en algún momento, fueron adoptadas (endosimbiosis) por otra célula primitiva de mayor tamańo (protobionte), proveniente de un hábitat diferente y cercano, que había perdido su pared celular. Este fenómeno promovió la formación de una célula más evolucionada, muy semejante a una eucariota que mejoró de este modo su chance de sobrevivir y reproducirse (De Duve, 1996:38-40). Margulis principal propugnadora de la teoría endosimbióntica seriada, aduce que los cloroplastos estaban contenidos en cianobacterias, y las mitocondrias en proteobacterias (Marguli, 1996: 1071-1076).
Las células (eucariotas), productos de esta unión tienen registros fósiles datados en 2000 Ma. El análisis evolutivo y analógico ha permitido consolidar la clasificación de las células primitivas en procariotas propiamente dichas, eubacterias y arqueobacterias, con DNA circular en mínima cantidad, inmerso en un nucleoide y el citoplasma, con proteínas pero sin histona; y eucariotas, con DNA constituído por proteínas e histonas, organizados en cromosomas, y rodeado de una membrana nuclear. De estas últimas han derivado: protistas, hongos, animales y plantas (Schopf, 1979:53-55, Knoll,1994:64-73).
Los eventos más importantes relacionados con la evolución de las procariotas pueden resumirse así: la frecuencia de cambios evolutivos en las procariotas era muy lenta (hipobraditelia), y seguían con buena fortuna la regla de la supervivencia del no especializado, es decir eran generalistas, por lo que quizás, predominaron en el Precámbrico (Schopf, 1994.6735). De otro lado, habían consolidado en sistema sencillo de metabolismo basado en la rotura de la molécula de glucosa, portadora de 6 átomos de carbono en dos, formado dos moléculas de piruvato (glucólisis anaeróbica). Dado que la concentración de oxígeno en esta época era oscilante o mínima, pero siempre por debajo del 50 % de las concentraciones actuales, sus sistemas metabólicos optaron por obviarlo, desenvolviéndose mayormente en medios anaeróbicos, con lo que la ganancia neta de energía fue de sólo dos moléculas de ATP (adenosintrifosfato) (Logan et al., 1995:53-56).
Pero el oxígeno liberado lentamente como subproducto de la fotosíntesis de procariotas ancestrales se había ido acumulando en la atmósfera, determinando un nuevo ciclo de adaptación biológica. Las bacterias que al principio apenas toleraban el oxígeno pronto lo utilizaron activamente (Knoll, 1991:64-73). Las eucariotas, ańadieron a la glucólisis anaeróbica reacciones dependientes de concentraciones mayores de oxígeno, como la reutilización del ácido cítrico, el mismo que partiendo de la misma molécula única de glucosa produjo finalmente agua, calor, CO2, y 34 moléculas de ATP. La edificación del ciclo del ácido cítrico como sistema casi único o universal para acceder a metas mayores permitió que los diversos sistemas metabólicos primitivos que fijaban nitrógeno, utilizaban el hidrógeno del H2S o elementos inorgánicos para generar energía; pasaran a un segundo plano (Knoll,1991:68-70). Demás está decir que las mayores concentraciones de oxígeno permitieron que la frecuencia evolutiva avanzara a niveles de normotelia, entre los 1700 - 1000 Ma.
Una amplia diversidad de eucariotas en forma de algas rojas, verdes y cromofíticas, es registrada recién desde hace 1000 Ma, a la fecha (Knoll, 1994:6743). El oxígeno permitió a las eucariotas mayor capacidad par formar esqueletos carbonados de enlaces covalentes, de hasta 40 uniones, necesarios para conformar ácidos grasos, esteroles: colesterol y hormonas esteroides en animales, y pigmentos en las plantas (Schopf, 1979:56-57).
| [Componente Mapa de imágenes de FrontPage] |
La evolución hacia funciones cada vez más complicadas promovió el incremento del tamańo de las células. Las procariotas raramente pasan de los 60 milimicrones. La aerobiosis permitió pasar de la simple división binaria de las PROCARIOTAS a la división celular por mitosis (reproducción sexual), mediante la cual las variaciones genéticas de los progenitores pasan a la descendencia, en forma de combinaciones nuevas permitiendo que las nuevas adaptaciones se extiendan rápidamente a través de una población acelerando la marcha evolutiva. Luego aparecerían las formas grandes y complejas, las que se diversificaron rápidamente, a inicios del Cámbrico (Knoll, 1994:6747-6750). Algunos sistemas metabólicos ancestrales (glucólisis anaeróbica) aún se hallan presentes en los sistemas pluricelurares eucariotas, y se actualizan en caso de insuficiencia del ciclo del ácido cítrico, cuando las exigencias de oxígeno superan a lo aportado por la sangre y los pulmones (Schopf, 1979:58-60).
Los capítulos 2 al 10 están basados en el trabajo del Dr. Victor Mechán Méndez de la UPCH
![]()
![]()
11.-La revolución darwiniana
Aunque la idea de la evolución tenía precedentes, no fue hasta 1859, con la aparición de la obra El origen de las especies del naturalista británico Charles Darwin, que la idea de la evolución se estableció definitivamente. Darwin recopiló e interpretó un gran número de observaciones y experimentos de muy diversas disciplinas de investigación y los presentó como un argumento irrefutable en favor del hecho de la evolución. Pero Darwin suministró además un mecanismo para explicar las adaptaciones complejas y características de los seres vivos: la selección natural. żQué significó la teoría de la evolución y de la selección natural en el contexto de la biología del siglo XIX? En 1802 el teólogo W. Paley publica la obra Teología natural, en donde arguye que el diseńo funcional de los organismos evidenciaba la existencia de un creador omnisapiente. Según él, el ojo humano, con su delicado diseńo, constituía una prueba concluyente de la existencia de Dios. Para los naturalistas que querían explicar los fenómenos biológicos por procesos naturales, explicar la adaptación, la maravillosa adecuación de los organismos a su ambiente, constituía el problema fundamental.
El gran reto de Darwin era explicar las complejas adaptaciones de los organismos vivos, como el diseńo funcional de un ojo, por mecanismos naturales. La solución de Darwin fue proponer el mecanismo de la selección natural
El argumento del diseńo de Paley tenía una gran influencia en los naturalistas del XIX, a pesar de que esta visión intervencionista violaba flagrantemente el concepto de naturaleza que se había establecido con el desarrollo de la física en los siglos XVI y XVII. Los fenómenos del Universo, según esta nueva concepción, eran explicables por procesos naturales. La naturaleza, per se, era un objeto lícito para preguntar y contestar científicamente. Con el Origen de Darwin se introduce esta revolución en la Biología. Lo verdaderamente revolucionario en Darwin fue el proponer un mecanismo natural para explicar la génesis, diversidad y adaptación de los organismos.
El naturalista británico Charles Darwin (1809-1882) introdujo en su libro El origen de las especies (1859) dos ideas revolucionarias: la evolución biológica y la selección natural
Para imponer su teoría de la evolución y de la selección natural, Darwin tuvo que introducir una nueva forma de entender la variación en la naturaleza, el pensamiento poblacional. En el tiempo de Darwin las especies se consideraban entidades fijas e inmutables; representaban a un tipo platónico, la idea perfecta de la mente de su creador. Las diferencias en la forma, en la conducta, o en la fisiología de los organismos de una especie no eran más que imperfecciones, errores en la materialización de la idea de la especie. En contraste con esta visión esencialista dominante, la variación individual, lejos de ser trivial, es para Darwin la piedra angular de la evolución. La variación en el seno de las especies o poblaciones es lo único real, es la materia prima de la evolución, a partir de la que se va a crear toda la diversidad biológica. Son las diferencias existentes entre los organismos de una especie las que, al magnificarse en el espacio y en el tiempo, producirán nuevas poblaciones, nuevas especies, y por extensión, toda la diversidad biológica.
Bajo la visión darwiniana, la variación es la única realidad de las especies. No hay un color de piel en la especie humana ideal o arquetípico. Cada individuo con su variación característica es un elemento esencial de nuestra especie
Los estudios y afirmaciones acerca de la evolución generalmente se refieren a uno de dos aspectos distintos: (1) las investigaciones acerca del hecho de la evolución y (2) las que se refieren al mecanismo de la evolución. Las primeras abarcan las disciplinas biológicas, tales como la paleontología, la clasificación, la biología comparada, la biología de poblaciones,... que muestran de manera inequívoca el hecho de la evolución. Las segundas, las afirmaciones acerca del mecanismo de la evolución, son las que nos informan de los factores, fuerzas o procesos que producen el cambio evolutivo, es decir, los mecanismos naturales que causan la descendencia con modificación. Una analogía cotidiana que ilustra esta distinción es la del tiempo meteorológico. Las precipitaciones, los vientos, las gotas frías, los tifones, son las evidencias que constituyen las afirmaciones de hecho del tiempo atmosférico. Ahora bien, si queremos explicar el porqué se dan los diferentes fenómenos meteorológicos, entonces nos tenemos que introducir en el ámbito de las afirmaciones del proceso o de los mecanismos meteorológicos. Debemos proponer los factores, tales como las diferencias de temperatura entre distintas masas de aire, que producen los fenómenos meteorológicos.
La evolución que se da en una escala reducida, en el interior de una especie y en el intervalo de unas pocas generaciones, se denomina microevolución. La macroevolución es la evolución a gran escala, y abarca periodos considerables de tiempo, y grandes procesos de transformación; en el caso más extremo comprendería toda la evolución de la vida. Se pueden efectuar experimentos y/o observaciones de poblaciones de especies actuales a pequeńa escala y obtener evidencia directa de evolución. Hay muchos ejemplos en los que se detecta la evolución en acción, como el clásico caso del melanismo industrial que se comentará más adelante. La selección artificial efectuada por el hombre en el perro o el caballo son también claros ejemplos que muestran el potencial de modificación de una especie. Por su propia dimensión temporal, no podemos demostrar la macroevolución directamente, exceptuando los casos de creación de nuevas especies de plantas mediante cruzamiento de especies distintas por el hombre. Aunque la evidencia evolutiva que suministra el registro fósil, la biología comparada, y la biología molecular es indirecta, no por ello es menos concluyente a la hora de demostrar la comunidad de origen de todos los organismos. Las diferentes razas de perro que el hombre ha obtenido por selección artificial ilustran el potencial de cambio que tienen las especies.
Los sedimentos que se han ido acumulando sobre la corteza de la tierra durante su historia geológica dejan una huella inestimable, generalmente en forma de huesos o esqueletos duros petrificados, de organismos muertos en el pasado: son los fósiles. El registro fósil es una ventana maravillosa a la historia de la vida. Si no existiera no podríamos inventarlo. Sin él, el vacío acerca de la evolución de la vida sobre la tierra sería insustituible. Podríamos especular, teorizar infinitamente, pero nunca dispondríamos de la evidencia con la que probar nuestras hipótesis más atrevidas. żQuien podría haber imaginado que la Tierra estuvo dominada durante 150 millones de ańos por unos reptiles inmensos y fantásticos, los dinosaurios, que desaparecieron en un instante relativo de tiempo, si no hubieran existido fósiles de dinosaurios que nos lo contasen? La desintegración de los elementos químicos radiactivos que hay en las rocas ha permitido estimar que la Tierra se originó alrededor de hace 4600 millones de ańos. La Tierra, que era una esfera caliente, se enfría gradualmente, iniciándose un periodo de evolución química que culminará con la formación de las primeras células. En Australia y África se han encontrado sedimentos retenidos y fijados por bacterias de hace 3600 millones de ańos, lo que hace que esta fecha sea una estima mínima de la edad de inicio de la evolución biológica. La magnitud del tiempo en el que ha transcurrido la evolución se escapa completamente a nuestra comprensión, no podemos siquiera imaginar, limitados a la minúscula escala de nuestro tiempo vital, el potencial de transformación que suponen 3600 millones de ańos de evolución.
El tiempo geológico se ha dividido en una serie de etapas jerárquicas, las eras, los períodos, y las épocas, que no siguen una cronología lineal, sino que es una crónica de los momentos claves de la historia de la vida. Las transiciones entre las cuatro eras, la Precámbrica, la Paleozoica, la Mesozoica y la Cenozoica representan grandes cambios en las fauna y flora de toda la Tierra. En el primer período de la era Paleozoica, el Cámbrico, hace 570 millones de ańos, aparecen de golpe en el registro fósil los animales pluricelulares que tienen partes duras, como las conchas, y exoesqueletos,... El final del Paleozoico coincide con la mayor extinción habida en la Tierra, en la que desaparecieron el 96% de las especies. Al final del Mesozoico, en la transición entre el período Cretácico y Terciario, se da la conocida extinción de los dinosaurios, junto a un 70% de las especies existentes.
![]()
![]()
12.-Cronología de los siete momentos estelares de la evolución
żQué nos enseńa el registro fósil sobre la historia de la vida sobre la Tierra? Esta es una lista de los acontecimientos más importantes
Origen de la célula procariota 3600 M (M=Millones de ańos) | |
Origen de la célula eucariota 1400 M | |
Origen de la fauna de animales pluricelulares 650 M | |
Fauna de la explosión cámbrica 570 M | |
Origen de los vertebrados terrestres 360 M | |
Extinción de los dinosaurios. La antorcha pasa de los dinosaurios a los mamíferos 65 M | |
Origen de Homo sapiens 0,1 M. |
Si toda la historia de la Tierra la comprimiésemos en una hora, a los 20 minutos aparecerían las bacterias, a los 55 los dinosaurios, los antropoides aparecen a 40 segundos antes del final, y los humanos al cumplirse la hora.
Hay una coherencia entre el registro fósil y el origen evolutivo de las especies y sus grupos correspondientes. Si la vida se debiera a la creación espontánea e independiente de especies, no tendríamos porqué esperar que el hombre surgiera con los mamíferos, podríamos haber surgido en cualquier momento, cuando aparecieron los peces, o los dinosaurios, o antes de que surgieran los primates.
![]()
![]()
13.-Las extinciones
La evolución no tiene dirección
La historia de la vida es una historia de extinciones y muerte,... con unos pocos supervivientes. El 99,9% de las especies que han existido alguna vez están hoy extintas. Grupos enteros de organismos, como los dinosaurios, los trilobites, los ammonoideos, se han extinguido para siempre sin dejar descendiente alguno.
Como seńala el reconocido paleontólogo S. Gould, el registro fósil no es un relato convencional que conduce a los diferentes linajes a más excelencia, más complejidad, más diversidad. La historia de la vida no muestra dirección ni sentido. La evolución es una narración de eliminación masiva seguida de diferenciación en el interior de unos cuantos supervivientes. Es a priori imposible determinar la dirección de la evolución porque la importancia de los acontecimientos concretos, contingentes, como la extinción o no de un grupo de organismos en el caso de una extinción en masa, o la posesión o no de una variante adaptativa adecuada cuando ésta es requerida, son los verdaderos agentes de la historia.
Hay dos regímenes de extinciones: la extinción normal, que afecta a las especies que dejan de seguir a su ambiente en su lucha diaria por adaptarse, y la extinción en masa, que son más rápidas y devastadoras en cuanto a su magnitud. Al menos han ocurrido cinco extinciones en masa, y han dejado muchos huecos ecológicos que han permitido que sean ocupados por los descendientes de las especies supervivientes. Esta ocupación del bioespacio disponible suele ir acompańada de una rápida y extensa diversificación morfológica que se denomina radiación adaptativa. No se sabe con certeza la causa de las extinciones masivas, aunque las causas físicas como el impacto de asteroides o el cambio climático parecen más probables que las biológicas. Según una hipótesis reciente, hay ciclos de extinción masiva aproximadamente cada 26 millones de ańos, y el impacto periódico de lluvias de cometas sobre la Tierra podría explicar dicho ciclo.
Si tuviéramos la oportunidad de volver a un momento del pasado, el período previo al Cámbrico sería sin duda uno de los más atractivos. En el Cámbrico, hace 570 M, se produce la gran explosión en el registro fósil de los primeros animales pluricelulares con partes duras. Darwin se preguntaba por qué estos primeros animales eran ya anatómicamente complejos y sin precursores aparentes. La respuesta está en el yacimiento de Burguess Shale, situado en las montańas rocosas canadienses. Debido a condiciones muy especiales de conservación, aquí se encuentra la única fauna de cuerpo blando (sin estructuras duras) que existe de un tiempo inmediatamente anterior a la explosión cámbrica.
La gran sorpresa de Burguess Shale es que las pocas especies analizadas contienen una disparidad de diseńos anatómicos que exceden, con mucho, la gama moderna que hay en todo el mundo. En ninguna fauna posterior se repite la riqueza anatómica de este yacimiento. De los 120 géneros analizados, 20 son diseńos de artrópodos únicos, y además de estar representados los 4 grupos de animales que hoy existen, hay 8 diseńos que no encajan en ningún grupo animal conocido. La época de Burguess Shale parece ser que fue un tiempo asombroso de experimentación, una era de gran flexibilidad evolutiva, que fue seguido de una gran extinción.
Opabinia: una vida extrańa de un remoto pasado
Opabinia fue el fósil de Burguess Shale que reveló la caja de sorpresas que había en este yacimiento. Su diseńo singular no pertenece a ningún grupo animal conocido. ˇTiene 5 ojos!, trompa flexible formada por un tubo cilíndrico estriado, un tubo digestivo en forma de U, un tronco de 15 segmentos, con lóbulos delgados laterales. Los 3 segmentos últimos forman una cola con tres pares de hojas delgadas y lobuladas dirigidas hacia afuera.
Opabinia fue el primer fósil del yacimiento de Burguess Shale que demostró la singularidad morfológica de las primeras formas pluricelulares. Tiene 5 ojos, trompa flexible, tubo digestivo en forma de U, un tronco de 15 segmentos, los 3 últimos forman una cola.
Durante el periodo Triásico surgen los dinosaurios de una línea de los reptiles e inician una dinastía sobre el medio terrestre que habría de durar 150 M. Se han identificado más de 350 especies de dinosaurios, y se cree que esta es una estima muy inferior a la de las especies que realmente existieron. Entre los dinosaurios están los animales más grandes que alguna vez hayan vivido sobre la Tierra. El Seismosaurus, el mayor herbívoro conocido, tenía unas 40 metros de longitud. El Tyrannosaurus rex, uno de los mayores carnívoros, tenía 12 m de longitud. Los dinosaurios se dividen en dos grandes órdenes, los de pelvis de lagarto (Saurisquios), que comprenden tanto especies carnívoras como herbívoras, y los de pelvis de ave (Ornistiquios), cuyas especies eran todas herbívoras. Los dinosaurios se extinguieron hace 65 M junto a otras muchas especies. Se ha propuesto que el impacto de un asteroide sobre la superficie terrestre podría ser la causa de la extinción en masa.
żQuien podría haber imaginado que la Tierra estuvo dominada durante 150 millones de ańos por unos reptiles inmensos y fantásticos, los dinosaurios, que desaparecieron en un instante relativo de tiempo, si no hubieran existido fósiles de dinosaurios que nos lo contasen?
La sexta extinción en masa
El destino natural de cada especie es su extinción. Pero la vida continúa porque muchas especies dejan especies descendientes antes de morir. La diversidad biológica es un proceso dinámico que resulta del equilibrio entre la extinción y la producción de especies. Ninguna de los dos puede predominar por mucho tiempo. Una extinción que fuese superior a la producción de especies durante muchas generaciones conduciría a la pérdida de la vida sobre la Tierra, mientras que una situación inversa llevaría al agotamiento de los recursos y por tanto a la extinción. Actualmente desaparece una especie cada 15 minutos. El crecimiento desmesurado de la especie humana ha aumentado en mil veces la tasa normal de extinción, creando una situación que es análoga a la de una gran catástrofe. Estamos frente a la sexta, y quizá definitiva, gran extinción de la Tierra.
![]()
![]()
14.-Biología comparada: homología y analogía
Cuando uno observa similitudes entre especies, se pueden distinguir entre dos tipos de semejanzas, la analogía y la homología. El ala de un ave y el de una mosca forman una extensión plana y tienen un movimiento de aleteo similar; los peces, los delfines, o los pingüinos tienen una sección transversal aplanada que les permite desplazarse por el agua. Estas semejanzas, llamadas analogías, son más bien superficiales y se deben a que estos organismos están sometidos a las mismas restricciones funcionales o adaptativas, y no son debidas a que posean un antepasado común reciente. Ejemplo de analogía. La forma aplanada de los vertebrados marinos es una adaptación al desplazamiento por agua.En contraste con la analogía, una homología es la similitud que hay entre caracteres de distintas especies debido a que tienen un origen común, y no a la acción directa de una presión funcional. Por ejemplo, todos los tetrápodos (animales vertebrados terrestres con cuatro extremidades) tienen una la extremidad de cinco dedos, y esta se encuentra tanto en las alas de la aves y de murciélagos como en la mano del ser humano, a pesar que estas extremidades representan unos papeles funcionales muy distintos. La razón de esta estructura común es que todos los tetrápodos conservamos la misma estructura básica de la especie ancestral original.
La homología es la base de la clasificación
La clasificación se basa en la comparación de los caracteres de las especies, y los caracteres homólogos son los elementos claves para establecer una clasificación evolutiva. Si las especies proceden de otras especies por evolución, y además no varían tan rápidamente como para perder toda su herencia histórica, se esperaría que los distintos seres vivos compartieran una serie de caracteres homólogos. El análisis de los diferentes caracteres fenotípicos, como la morfología, la conducta, los cromosomas, la anatomía externa e interna, el desarrollo embrionario, el metabolismo, la variación genética y proteica muestran que las especies presentan semejanzas homológas en todos los niveles del fenotipo. Cuanto más próxima sean la especies, mayor será el grado de semejanza, y lo contrario también es cierto, cuanto más alejada estén menos semejanzas encontraremos. Así, las diferencias que hoy vemos entre las especies se deben a las nuevas variaciones que han adquirido desde su separación del antepasado común. Las similitudes que atribuimos a las homologías no podrían explicarse si las especies se originasen independientemente unas de otras. Homología: todos los tetrápodos tienen una extremidad con cinco dedos, aunque tengan diferentes funciones.
Un caso especialmente significativo de homología es la de los órganos vestigiales o residuales. La pelvis de los tetrápodos es una estructura ósea cuya función es articular las extremidades posteriores. Las ballenas son tetrápodos cuyos antepasados mamíferos abandonaron la tierra para habitar en el mar. En esta nuevo medio las ballenas perdieron sus extremidades inferiores, pero aún conservan como huella acusatoria de su pasado tetrápodo, la serie completa de los huesos de la pelvis. De forma similar, las serpientes presentan vestigios de la extremidad posterior que portaban sus antepasados. La reducida pelvis de los cetáceos es un órgano vestigial que prueba su origen tetrápodo.
Biología molecular y homología
La biología molecular ha suministrado la evidencia más universal de homología. Todos los organismos vivos compartimos el mismo material hereditario, el ADN, una molécula helicoidal cuya información se encuentra codificada en 4 letras o nucleótidos distintos. Igualmente, el código genético es Universal, todos los organismos comparten el mismo diccionario que da el significado a la secuencia de DNA. Ambos ejemplos son pruebas muy robustas de la relación íntima que existe en todo lo vivo. El ADN es una molécula helicoidal que tiene información genética codificada a partir de cuatro letras o nucleótidos distintos.
![]()
![]()
15.-El árbol de la vida: filogenia
Si la historia de la vida es cambio y ramificación por descendencia, entonces su representación sería la de un árbol o filogenia, en la que el tronco y las ramas internas se corresponderían a los antepasados de las especies actuales y los extremos de las ramas externas serían las especies actuales. żComo se establece una filogenia? Ordenando las especies actuales atendiendo a la similaridad morfológica y/o genética de sus caracteres homólogos.
El botánico sueco Carolus Linnaeus (1707-78) ideó el sistema de clasificación que se utiliza, con algunas modificaciones, hoy en día. Hay siete niveles inclusivos de clasificación, que son, de menor a mayor, la especie, el género, la familia, el orden, la clase, el tipo o Phylum y el reino. El nombre científico de cada especie tiene dos partes, el león, por ejemplo, se denomina Panthera leo. La primera parte se refiere al género y la segunda a la especie. Consideremos un ejemplo de como se agrupan las especies actuales en las distintas categorías linneanas. El león, la pantera, el tigre, pertenecen al género Panthera, que junto al género Felix (el gato doméstico) y otros se agrupan en la familia de los felinos. Los felinos, con los cánidos y úrsidos, constituyen el orden de los carnívoros. Primates, roedores, carnívoros, ... se reúnen en la clase mamíferos. Estos organismos comparten características como el amamantar a sus crías con leche, gestarlas en el útero mediante un órgano complejo, la placenta. Su piel está protegida por pelaje o pelos. Mamíferos, aves, reptiles, anfibios, y peces se reúnen en un solo tipo o phylum, porque todos tienen columna vertebral, un máximo de cuatro miembros, y sangre roja con hemoglobina, son los cordados. Los insectos, las arańas, los crustáceos, y los ciempiés, se clasifican en otro tipo, los artrópodos. Las almejas, caracoles y calamares se agrupan en los moluscos, y así sucesivamente. El tipo cordado, artrópodo, molusco y otros forman el reino Animal. El árbol de la vida se ordena siguiendo divisiones que van de características generales a aspectos más específicos. El botánico sueco Carolus Linnaeus (1707-78) creó el sistema de clasificación que se utiliza hoy en día.
Significado de los niveles jerárquicos de clasificación
żPor qué se ordena la diversidad biológica en diferentes niveles jerárquicos? Los rasgos de las divisiones más generales corresponderían a adaptaciones básicas o principales que surgieron en los momentos iniciales de la evolución de los especies progenitoras de estos grupos. Por ejemplo, hay cinco grandes reinos, las moneras, los protistas, los hongos, las plantas y los animales, que se corresponden con las cinco diferenciaciones principales de la vida sobre la Tierra. El Phylum o tipo es la siguiente unidad básica de diferenciación entre reinos, y podrían entenderse como proyectos básicos fundamentales de anatomía. En el reino Animal estos planos fundamentales serían las esponjas, los anélidos (gusanos), los artrópodos (insectos, crustáceos,...), los cordados, .... A partir de estas adaptaciones principales surgen posteriormente nuevas variaciones o subadaptaciones diversas, que por tener un menor tiempo de evolución, tienen un rango clasificatorio menor.
La clasificación de la diversidad biológica es una prueba de la evolución
El hecho que la diversidad de la vida esté jerarquizada es un fuerte argumento en favor de la evolución. El que las especies compartan estructuras anatómicas y adaptaciones básicas puede explicarse fácilmente si suponemos que las especies actuales compartieron antepasados, pero no esperaríamos dicho patrón si las especies hubieran sido creadas independientemente.
La universalidad de la molécula portadora de la información genética hace que el ADN sea un carácter muy apropiado para el estudio comparativo y filogenético de las especies. Morfológicamente no es posible comparar una bacteria con un hombre, sin embargo si que es posible establecer una comparación con moléculas de ADN de ambos organismos, ya que están formadas por el mismo lenguaje de bases. Con datos de secuencias podemos comparar cualesquier grupo de organismos, por distantes que sean. Los datos moleculares tienen otras propiedades adicionales que todas juntas los convierten en el carácter ideal de estudios filogenéticos. Muchos trabajos obtienen y analizan las secuencias de genes y proteínas de diferentes especies para resolver cuestiones todavía dudosas de relaciones entre organismos. Los datos moleculares han demostrado que nuestra especie está mucho más cerca del chimpancé y el gorila de lo que creíamos.
El análisis molecular de secuencias también nos ha enseńado que hay una división en la raíz misma del árbol de la vida que es más fundamental que la división de 5 reinos que se enseńa normalmente. En lugar de los dos tipos celulares canónicos, los procariotas y eucariotas, hay tres tipos principales de células, las arqueobacterias, las eubacterias y los eucariotas. Este nuevo árbol recibe el nombre de árbol filogenético universal.
![]()
![]()
16.-Evolución humana
Las semejanzas morfológicas, bioquímicas, y genéticas sitúan al ser humano en el orden de los primates de la clase mamíferos. Dentro de los primates, son el chimpancé, el gorila y el orangután, sus parientes más próximos. Los datos de comparación de secuencias muestran que hay una similitud del 98,5% entre el ADN humano y el del chimpancé. Esta semejanza es mayor que la que existe entre el chimpancé y el gorila o el gorila y nosotros, por lo que el chimpancé y los humanos compartimos un antecesor común más reciente que ambos con los gorilas. Esta cercanía, que se ha estimado en 5M de ańos, es mucho mayor de lo que se había inferido sólo con datos morfológicos, y muestra la capacidad de los datos de ADN para desvelar relaciones de parentesco. En la evolución humana existen dos grandes adquisiciones, la marcha bípeda, y el desarrollo extraordinario del cerebro. El registro fósil nos muestra que la postura erguida precedió al desarrollo cerebral y que África es la cuna de la humanidad. El Australopithecus, de una antigüedad de 1,5-5M de ańos es el primer mono antropoide de marcha bípeda. Su capacidad craneal era similar a la del chimpancé y gorila actual. El Homo habilis y el Homo erectus son las líneas que siguen cronológicamente hasta la llegada de nuestra especie, Homo sapiens, hace 100.000 ańos.
En 1997 se publica la secuencia de un trozo del ADN del primer fósil que se encontró del hombre de Neandertal, una subespecie extinta de la especie humana. Es la primera vez que se obtiene la secuencia molecular de un fósil humano. Cuando la secuencia se comparó con secuencias homólogas de ADN humano actual se dedujo que el antepasado común de nosotros y el hombre de Neandertal vivió hace 500.000 ańos, de lo que se concluye que el hombre de Neandertal se extinguió sin mezclarse con el hombre actual. El análisis reciente de un trecho de la secuencia de ADN del hombre de Neandertal indica que esta subespecie humana no se mezcló con nosotros.
El futuro de la evolución humana
La evolución cultural humana no sigue las leyes de la selección natural y la herencia genética. Sigue un proceso de transmisión horizontal (entre individuos de una generación) y vertical (entre generaciones) que es mucho más veloz que los procesos de evolución biológica típicos. El ser humano está en el umbral de poder dirigir la evolución en la dirección que el crea conveniente. Podrá limitar enormemente los azares de la mutación y de la segregación genética y unión de gametos. Genes mutados que causan graves enfermedades a la especie humana podrán ser sustituidos por sus contrapartidas no deletéreas,... Pero paradójicamente la humanidad se enfrenta hoy a los retos de su propio éxito evolutivo. El crecimiento explosivo de su población, con las necesidades de espacio y recursos que genera, provoca la eliminación o reducción hasta tamańos insoportables de los hábitats de las especies. La tasa de extinción actual no es sostenible. Es posible que el ser humano pueda mantener la diversidad biológica en forma de semillas o células congeladas, e incluso que pueda compensar la pérdida de especies con la creación de nuevas mediante ingeniería genética. Pero la reducción drástica de espacios naturales limitará inevitablemente la diversidad. Los grandes retos de la humanidad son el control del crecimiento de la población, la eliminación de las desigualdades socioeconómicas, el mantenimiento de un desarrollo sostenido y viable, la conservación de hábitats naturales y de especies, y el desarrollo de una conciencia ética universal basada en el respeto a la diversidad de pueblos y culturas, y de todas las formas orgánicas que nos han acompańado, desde esa primera célula antecesora común, en este fascinante periplo singular e irrepetible que es nuestra historia biológica. La humanidad pronto podrá substituir genes mutados deletéreos por sus contrapartidas sanas
![]()
![]()
Se agregarán nuevos capítulos
.